

Changes in Average Trophic Level of Marine Consumers
Background
The MTL of marine organisms is an ecological indicator that reflects mainly the effects of fishing pressure on the structure of food webs. The indicator is based on biomass data and trophic level (TL) of the species. The TL, determined by predator-prey interactions, expresses the position of an organism within a food web. The lowest trophic level values (TL = 1) are assigned to the primary producers at the base of the food web, with herbivorous in the subsequent level (TL= 2) and the highest values (TL ≥ 4) are assigned to predators at the top. Thus, energy is transferred from the lower TLs to the higher ones following the consumer-resource connections of organisms in the food web. TL can be estimated using data from stomach content analyses and/or stable isotope analyses.
Most fisheries target species at a high TL (correlated with the largest sizes) and thus drive a decrease in the availability of predators within the ecosystem. This in turn forces fisheries into targeting species at ever lower TLs and thus a decline in the MTL of fisheries landings is commonly observed. Hypothetically, in those fisheries targeting small pelagics, the decrease of food availability to upper trophic levels could be reflected in downward trends of the indicator.
Traditionally, the indicator has been calculated using long historical series at large scale (regionally). However, recent studies showed the importance of analysing the indicator at local level showing highly impacted areas. Significant negative relationships between bottom trawling and MTL at small-scale spatial resolution were detected off the north Iberian coast (Preciado et al., 2019), despite the apparent recovery of top predators observed at a regional scale (Arroyo et al., 2019). The strong local influence of this anthropogenic pressure on bentho-demersal food webs showed the relevance of using local spatial resolution when investigating the effects of spatially heterogeneous pressures, such as fishing.
 decline it indicates that large fish are no longer available because fisheries have depleted top predators’ stocks.")
Figure 1: Diagram showing the rationale behind the MTL indicator: if Trophic Levels (TLs) decline it indicates that large fish are no longer available because fisheries have depleted top predators’ stocks.
Marine ecosystem functioning strongly depends on its structure, diversity and integrity, which are subjected to many environmental and anthropogenic pressures that cause alterations on ecosystem components at high, intermediate and low trophic levels (Cury et al., 2005; Bundy et al., 2010; Link et al., 2010; Heymans et al., 2014; Coll et al., 2016). Food webs, also called trophic networks, represent the most elemental structure of the ecosystem, illustrated by the consumer-resource interactions among organisms and evidencing the main pathways of energy flow through ecosystems, from primary producers to top predators. The knowledge of the functioning of these networks is key for understanding marine ecosystem functioning. Each life stage of an organism is associated with a particular trophic level and the energy is transferred from the lower trophic levels to the higher trophic levels following the interconnections between organisms within the food web.
The TL of a given species can be estimated using: i) dietary analysis from stomach contents, ii) stable isotopes analysis, and/or iii) modelling. Using stomach contents analyses, the TL of a species can be estimated by the TL of each prey category weighted by their relative abundance in its diet. Alternatively, stable isotope analyses rely on the ratio of 15N to 14N (δ15N) in the organisms’ tissues, as isotopic nitrogen increases regularly with each trophic step along the food webs.
The conceptual development of the MTL indicator, by Pauly et al., (1998), is known as ‘fishing down marine food webs’. This indicator proved that a decline in the MTL of the global fisheries catch during the 1990s revealed an erosion of the high trophic levels in the ecosystems worldwide, forcing fisheries to shift their pressure from the top predators to lower trophic levels, due to the decline in biomass of high trophic level species (Figure 1). Thus, this indicator aimed at highlighting the unsustainable fisheries practices of past decades. The lack of trends in this indicator does not necessarily indicate no impact, as highly impacted ecosystems might become so resilient that no trend means resilience. The indicator was adopted by the Conference of the Parties to the Convention on Biological Diversity as one of eight indicators to monitor achievement of a significant reduction in the rate of biodiversity loss by 2010.
Since then, the use of MTL has been further explored and additional hypotheses have emerged to explain the patterns observed in the MTL. The ‘fishing down food webs’ concept as defined above was actually detected in the North Atlantic. Other scenarios were described in other parts of the world (Essington et al., 2006; Morato et al., 2006; Swartz et al., 2010; Stergiou and Tsikliras, 2011; Shannon et al., 2014) including for example the concept of fishing through the food webs (sequential addition of low trophic level species in fisheries); fisheries expansion (into deeper and further offshore waters with the inclusion of new high trophic level species); and fishing up the food webs (addition of new previously unfished high trophic level species).
Within the scientific community, it is considered that food web status should be based on ecosystem indicators which represent the ecosystem as whole, including as many species as possible and making sure different trophic levels are well represented. The Mean Trophic Level (MTL) indicator meets this requirement because it takes into account food web structure and uses a metric (the TL) that reflects energy transfer between predators and prey. MTL can be calculated using commercial landings or biomass estimates from scientific surveys, in combination with the estimated TL of the species. In this assessment, the indicator has been calculated taking into consideration as many taxonomic and functional groups of species as possible (benthic invertebrates, cephalopods, elasmobranchs, fish). Since groundfish trawl surveys target soft-bottom shelf habitats, the present assessment is primarily focused on bentho-demersal communities of the continental shelves of OSPAR Region IV and each sub-division. The methodological approach may however be transferred to other OSPAR Regions whenever adequate/appropriate data become available.
To compute the MTL indicator, landings have traditionally been the only source of biomass data, raising many concerns around their suitability for describing the ecosystem as a whole, due to the strong discrepancies between biomass caught and biomass landed among other shortcomings and uncertainties. For example, landings data are incomplete (i.e., they do not take into account illegal, unregulated and unreported species, or by-catch), and are subject to multiple biases induced by fisheries drivers (Branch et al., 2010; Pauly et al., 2013). In contrast, data from scientific surveys reflect actual changes in community composition because they are not influenced by market forces (i.e., selectively fishing for particular commercial species only) and include all species that fisheries by-catch and are commonly not reported with the landings. Although data coming from groundfish surveys have particular shortcomings (they are usually limited to a specific season, limited by the meshsize and underrepresenting the smallest species, and sometimes focus on commercial fish and cephalopods), the standardisation of IBT surveys together with the long-term series available online make scientific surveys the best data source to assess the MTL at the moment.
Estimates of the TL values of most demersal species are currently available in online databases and repositories (e.g., www.fishbase.org). However, these values might refer to specific populations and sampling times or be calculated as worldwide averages based on TL data from different ecosystems. In many cases these values do not reflect the characteristics of a given region where trophic interactions between species can be region-specific. Environmental or human-induced pressures can trigger changes in prey availability and predator selectivity, in turn causing variability in the TL values for a given species in space and time or even along ontogeny (Pinnegar et al., 2002; Chassot et al., 2008; Vinagre et al., 2012). Therefore, TL values need to be region-specific in order to increase the accuracy of the analyses and better represent feeding interactions in the study region.
In the OSPAR Intermediate Assessment 2017, the assessment of this indicator was performed for OSPAR Region IV (French and North Iberian continental shelves) based on scientific surveys and landings, and including different scenarios: MTL2,0 (including all consumers), MTL3,25 (equal to the MTI defined by Pauly and Watson in 2005), MTL4,0 (focus on higher predators) (https://oap.ospar.org/en/ospar-assessments/intermediate-assessment-2017/biodiversity-status/fish-and-food-webs/mtl-bay-biscay/). The main results showed no apparent change in the overall food web structure for MTL2,0, whereas a subtle increase in MTL was detected when focusing on the predator community (MTL3,25 and MTL4,0) associated with signs of recovery in these ecosystems (Arroyo et al., 2019).
The purpose of the present assessment, is to expand the analysis to other sub-divisions (Portuguese shelf and Gulf of Cadiz) and extend the historical series until 2021, as well as to analyse whether those regional-scale trends identified in 2017 can also be acknowledged at local level (haul/square). For that purpose, a spatio-temporal approach is performed, calculating the MTL in each haul by year and analysing the trend at the local scale. A case study in the North Iberian coast based on this approach has recently being published with promising results (Preciado et al., 2019), despite using a shorter time series. In addition to the expansion of the analyses in space (other sub-divisions) and time (2000-2021), MTL trends are also calculated in different scenarios, in order to identify potential changes of the trends with depth (i.e., coastal, shelf and deep ecosystems).
Data sources
Biomass data of species can be downloaded directly from: DATRAS in exchange format https://datras.ices.dk/Data_products/Download/Download_Data_public.aspx. In the present assessment, the indicator has been calculated starting in 2000 and using only data from the fourth quarter of the year (Q4) in order to keep consistency across the four sub-divisions under study. The differences in the DATRAS database uploaded by each country should be taken into account when calculating the indicator, as they may prevent comparisons across regions. In Region IV this was not an issue since one survey is carried out per country in each sub-division. There is also agreement on the type of gear used. The set of species reported are also crucial for the comparability of the indicator among sub-divisions, as the inclusion of some species or others (commercial versus non-commercial fish and cephalopods, and/or benthic invertebrates) can lead to erroneous results of the indicator or even to not being able to calculate it.
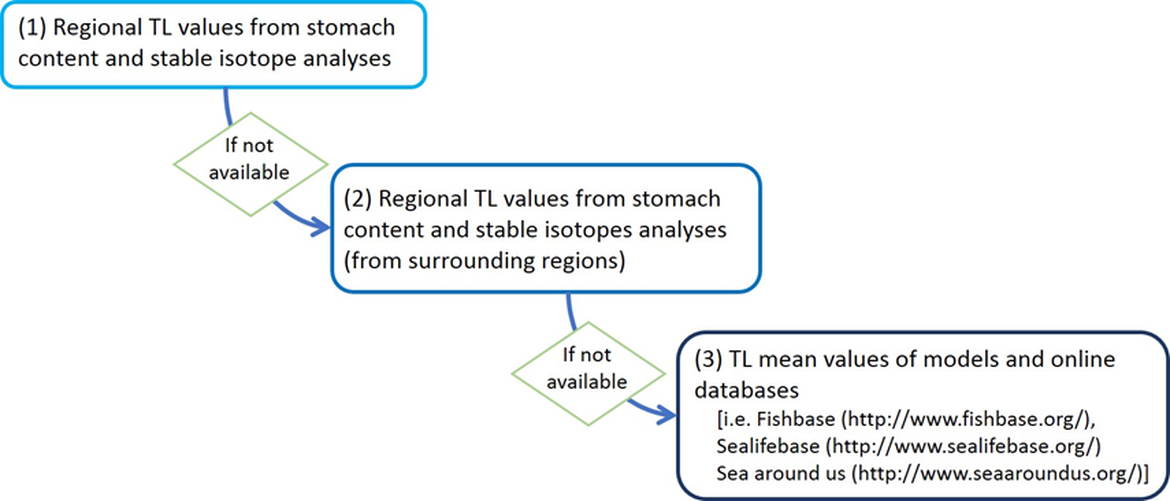
The TL estimated from stomach content analysis, stable isotope analysis or models can be collected from the literature for regional and non-regional areas. Otherwise, TLs are available in online databases [i.e. Fishbase (http://www.fishbase.org/), Sealifebase (http://www.sealifebase.org/) and Sea Around Us (http://www.seaaroundus.org/)]. For this assessment, information on TLs per species have been estimated by sub-division using local stomach contents and stable isotopes (e.g. Chouvelon et al., 2012; Lasalle et al., 2011; Lasalle et al., 2014; Le Loc’h et al., 2008), except for Portuguese waters where TL coming from the North Iberian shelf has been applied in the calculations. The TL databases by sub-division were compiled into a single database and populated by each country. This TL database for OSPAR Region IV is now available at the ODIMS repository (https://odims.ospar.org/en/submissions/ospar_trophic_level_2021_05/).
During the 2017 assessment, a preliminary work was performed in to collate regional TL estimations per species for OSPAR Region IV. However, the use of TL databases by sub-division was recommended in order to capture local trophic interactions and regional differences in the structure of the trophic networks. TL estimates by sub-division are then more relevant to calculate the MTL of such particular sub-division. The TL was compiled in three different steps, prioritising in each case local estimates over TL estimated at larger scale, and making use of the latter only when the former were unavailable, as shown in the following scheme:
To do this, only taxons at high taxonomic resolution (species, genus or family) were considered, excluding taxa identified at low taxonomic resolution (i.e., phylum, sub-phylum, class, superorder, order, suborder and infraorder). Nevertheless, in terms of biomass, taxa identified with low taxonomic resolution were practically negligible.
Uncertainty exists for each TL value estimated by diet studies, which is related to the spatio-temporal variability of the diet and differences of diet throughout the ontogeny (Pinnegar et al., 2002; Chassot et al., 2008; Vinagre et al., 2012). This uncertainty needs to be reported as a standard error for each TL value of a species.
Ecological and geographical scenarios
Different scenarios were considered for the calculation of MTL and MTL changes across time. Firstly, the MTL of the four sub-divisions was calculated independently, using the same methodology (specified in the following section) but based on local TL and biomass data.
For each of the sub-divisions, three different bathymetric ranges were considered, assessing independently MTL at the coastal (<30 m depth), continental shelf (30-200 m depth) and continental slope (>200 m depth). These bathymetric ranges reflect the fact that communities on regional seas are commonly structured along the depth gradient, and the community inhabiting the coastal area is fundamentally different from that inhabiting the deeper area, although both can be summarised into an MTL value. Thus, the bathymetric segregation allowed us to explore temporal changes in these communities independently, with the potential to identify divergent trends along the depth gradient. The coastal area (<30 m) did not have a sufficient sampling coverage in the North-western Iberian and Portuguese regional seas to be included in this assessment, so in these two sub-divisions only the continental shelf and slope were assessed.
Furthermore, for each sub-division and depth range three ecological cut-offs in the trophic structure of the community were considered. MTL2 included all species in the community except for primary producers. MTL3,25 included only those species with TL≥3,25, which are essentially secondary consumers and top predators, here called meso + top predators. Lastly, MTL4 included only species in the top of the food web, here called top predators (Figure a). Differentiating the ecological community in these thresholds makes it possible to identify patterns affecting only top predators (as might be the case of the “fishing down” hypothesis) which might be obscured by the high biomass of low trophic level species otherwise.
Finally, considering the fact that bottom trawl surveys do not specifically target pelagic species but are designed to capture and evaluate bentho-demersal species and that the biomass of pelagic species dominates the community showing very high variability, the bathymetric range and MTL cut-offs, including and excluding pelagic species, were considered in each sub-division. The combination of this geographic and ecological constrains resulted in 18 scenarios which are summarised in Table a.
| Top predators (MTL_4,0) | Meso + top predators (MTL_3,25) | All community (MTL_2,0) | ||
|---|---|---|---|---|
| Coastal (depth <30 m) | with pelagics | top predators | mesopredators + top predators | all consumers |
| without pelagics | top predators (excluding pelagic species) | Mesopredators + Top predators (excluding pelagic species) | all consumers (excluding pelagic species) | |
| Shelf (depth 30- 200 m) | with pelagics | top predators | mesopredators + top predators | all consumers |
| without pelagics | top predators (excluding pelagic species) | Mesopredators + Top predators (excluding pelagic species) | all consumers (excluding pelagic species) | |
| Deep (depth >200 m) | with pelagics | top predators | mesopredators + top predators | all consumers |
| without pelagics | top predators (excluding pelagic species) | Mesopredators + Top predators (excluding pelagic species) | all consumers (excluding pelagic species) |
Calculation of the Mean Trophic Level
Before performing the calculations and in order to select a set of species that have been consistently well sampled through the historical series, cumulative curves were applied to the biomass database to make a selection of species. Filter criteria should be agreed among scientists so that the assessment is consistent across regions. The following criteria were finally decided, based on the inflection points of species frequency of occurrence by space and time, and their abundance distribution:
- species that appeared at least in 50% of the years
- species that appeared at least in 10% of the hauls each year
- species that displayed an abundance above the quantile 10
The TL value and biomass of each species in each haul were used to calculate the MTL per haul. The indicator is then calculated for the whole community (all community, MTL2,0), including only species with TL higher than 3,25 (meso + top predators, MTL3,25) and including only species with TL higher than 4 (top predators, MTL4,0) (Figure a).
 all community (MTL_2,0), ii) mesopredators and top predators (MTL_3,25), iii) top predators (MTL_4,0).")
Figure a: Schematic diagram showing the composition of species/taxa included in each TL cut-off: i) all community (MTL_2,0), ii) mesopredators and top predators (MTL_3,25), iii) top predators (MTL_4,0).
The Mean Trophic Level indicator for each haul h can be calculated using the biomass and TL of species with the following formula:
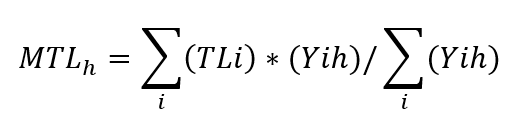
MTLh refers to the MTL indicator by haul
TLi refers to the trophic level estimation of species (group) i
Yih refers to the biomass (Y) of species (group) i in each haul h.
The resulting MTL values were explored to identify changes along the historical series in the different scenarios, by modelling the MTL as a function of year using Linear Models (LM) and/or Generalised Additive Models (GAM). All statistical analyses were performed using R software (R Core Team, 2022).
For the spatio-temporal approach, the calculation process follows the next steps for each of the scenarios (Figure b):
- Step 1. Calculate the mean MTL on a 0.2 x 0.2 degree grid over the whole time series.
- Step 2. For those grid cells having a sufficient number of data (n>10) assess any linear trend in MTL by year, reporting the slope of the linear model and its p-value.
- Step 3. Plot these results spatially, with MTL and the slope of MTL represented by a colour scale, and any significant trend represented by the cell of interest framed in bold.

Figure b: Testing framework for the calculation of the spatio-temporal approach: MTL by haul and by year
Assessment criteria
The assessment of the MTL is based on its trend analysis. To summarise the results of different scenarios and show a comprehensive message to the scientific and non-scientific community, the results of the trend analyses were binned into categories after McQuatters-Gollop et al., 2022. Colours are the result of the linear models showing increase/no-trend/decrease of the indicator. Trends were considered to be increasing or decreasing only when linear models were significant (*p<0.05, **p<0.01, ***p<0.001). The resulting categories are shown in Table b. Please note that the present assessment shows changes in the indicator starting in 2000 when the bentho-demersal ecosystems were already overexploited, so the values at the start of the series do not represent ecological target values. Any positive trend in MTL is therefore considered as a sign of recovery, while a non-significant trend can be considered as a sign of the high resilience characterising systems enduring high and sustained impacts.
Table b: Biodiversity status categories and colours used for the interpretation of the results. Trend analysis (linear model and significant tests) and expert judgement are used to set colours based on the categories proposed by McQuatters-Gollop et al., 2022.
| Biodiversity status categories | |
| Indicator value is below assessment threshold, or change in indicator represents a declining state, or indicator change is linked to increasing impact of anthropogenic pressures (including climate change), or indicator shows no change but state is considered unsatisfactory | |
| No assessment threshold and/or unclear if change represents declining or improving state, or indicator shows no change but uncertain if state represented is satisfactory | |
| Indicator value is above assessment threshold, or indicator represents improving state, or indicator shows no change but state is satisfactory | |
| Indicator was not assessed in a region due to lack of data, lack of expert resource, or lack of policy support. | |
Results
The Mean Trophic Level (MTL) indicator was assessed over a period of 19-22 years depending on the sub-division (OSPAR Region IV): French shelf (22 y), North Iberian shelf (22 y), Portuguese shelf (19 y) and Gulf of Cadiz shelf (21 y) (Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8) using biomass survey data and TL values specific for each sub-division. Above a TL of 4.0 (top predators) includes higher level predators of the bentho-demersal communities; above a TL of 3.25 (meso- and top predators) includes mid and upper trophic level fish, elasmobranchs and cephalopods; and above a TL of 2.0 (all community) includes all consumers.
Results on the temporal approach showed different trends in the four sub-divisions (Figure 2, Figure 3, Figure 4, Figure 5 and Table 1). The results observed in the French continental shelf indicate an overall increase of top predators in coastal and shelf ecosystem and no changes in deeper areas. General increasing trends were also detected when analysing the meso- and top predators. In the North Iberian coast contrasting results were detected with a decrease in the MTL of top predators while a significant increase of meso- top predators in all scenarios. In Portuguese waters a significant decrease of top predators in shelf ecosystems was apparent when including pelagics. In the Gulf of Cadiz, coastal ecosystems show decreasing trends when considering all community and meso + top predators. In contrast, shelf and deep ecosystems show signs of recovery at all trophic levels.
, depth ranges (coast, shelf, deep), with/without pelagic species.")
Figure 2: French continental shelf, groundfish survey data analyses. Trends in the MTL indicator with 18 different scenarios: TL cut-offs (top predators, meso + top predators, all community), depth ranges (coast, shelf, deep), with/without pelagic species.
, depth ranges (shelf, deep), with/without pelagic species.")
Figure 4: Portuguese continental shelf, groundfish survey data analyses. Trends in the MTL indicator with 12 different scenarios: TL cut-offs (top predators, meso + top predators, all community), depth ranges (shelf, deep), with/without pelagic species.
, depth ranges (shelf, deep), with/without pelagic species.")
Figure 3: North Iberian coast, groundfish survey data analyses. Trends in the MTL indicator with 12 different scenarios: TL cut-offs (top predators, meso + top predators, all community), depth ranges (shelf, deep), with/without pelagic species.
, depth ranges (coast, shelf, deep), with/without pelagic species.")
Figure 5: Gulf of Cadiz continental shelf, groundfish survey data analyses. Trends in the MTL indicator with 18 different scenarios: TL cut-offs (top predators, meso + top predators, all community), depth ranges (coast, shelf, deep), with/without pelagic species.
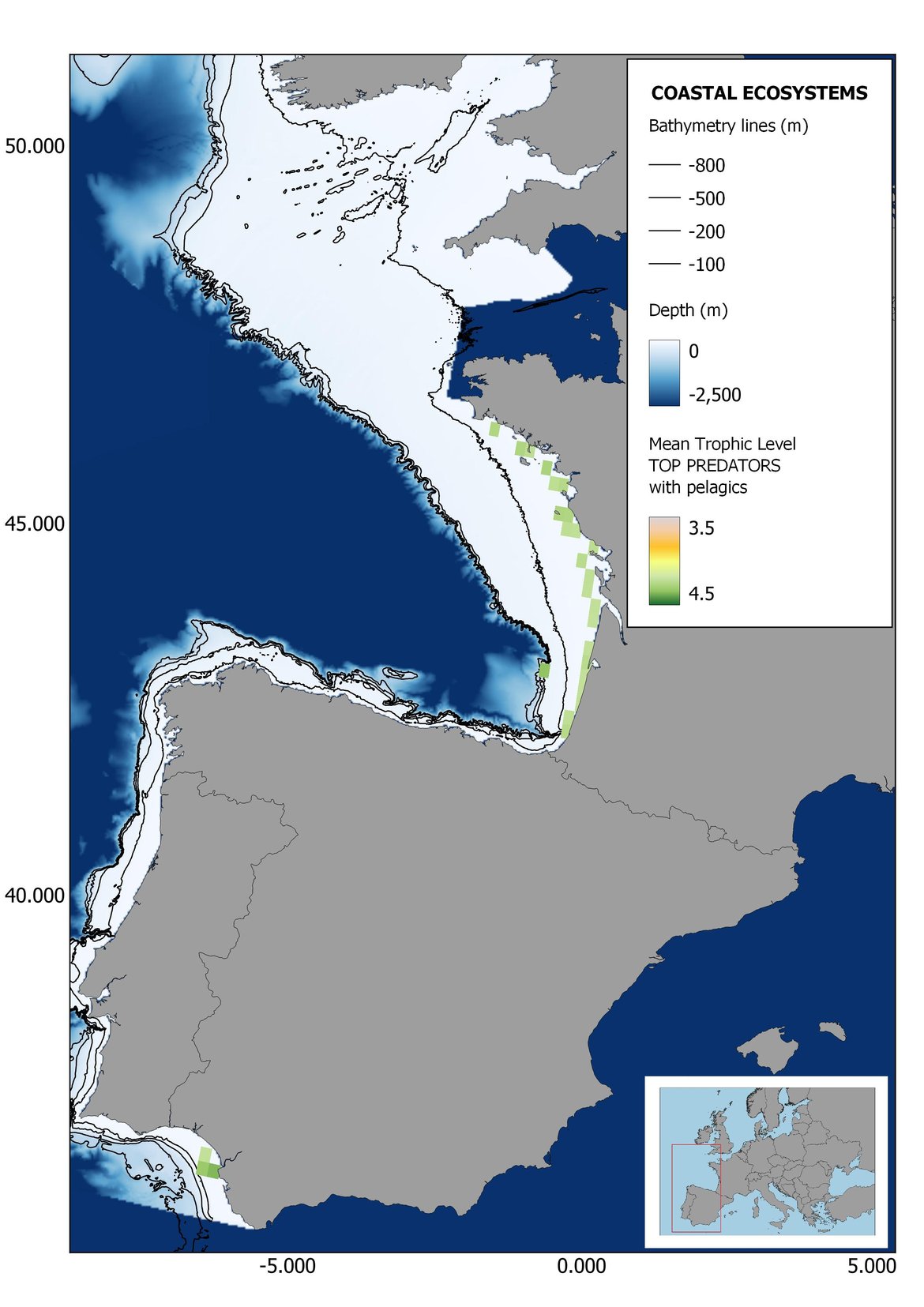
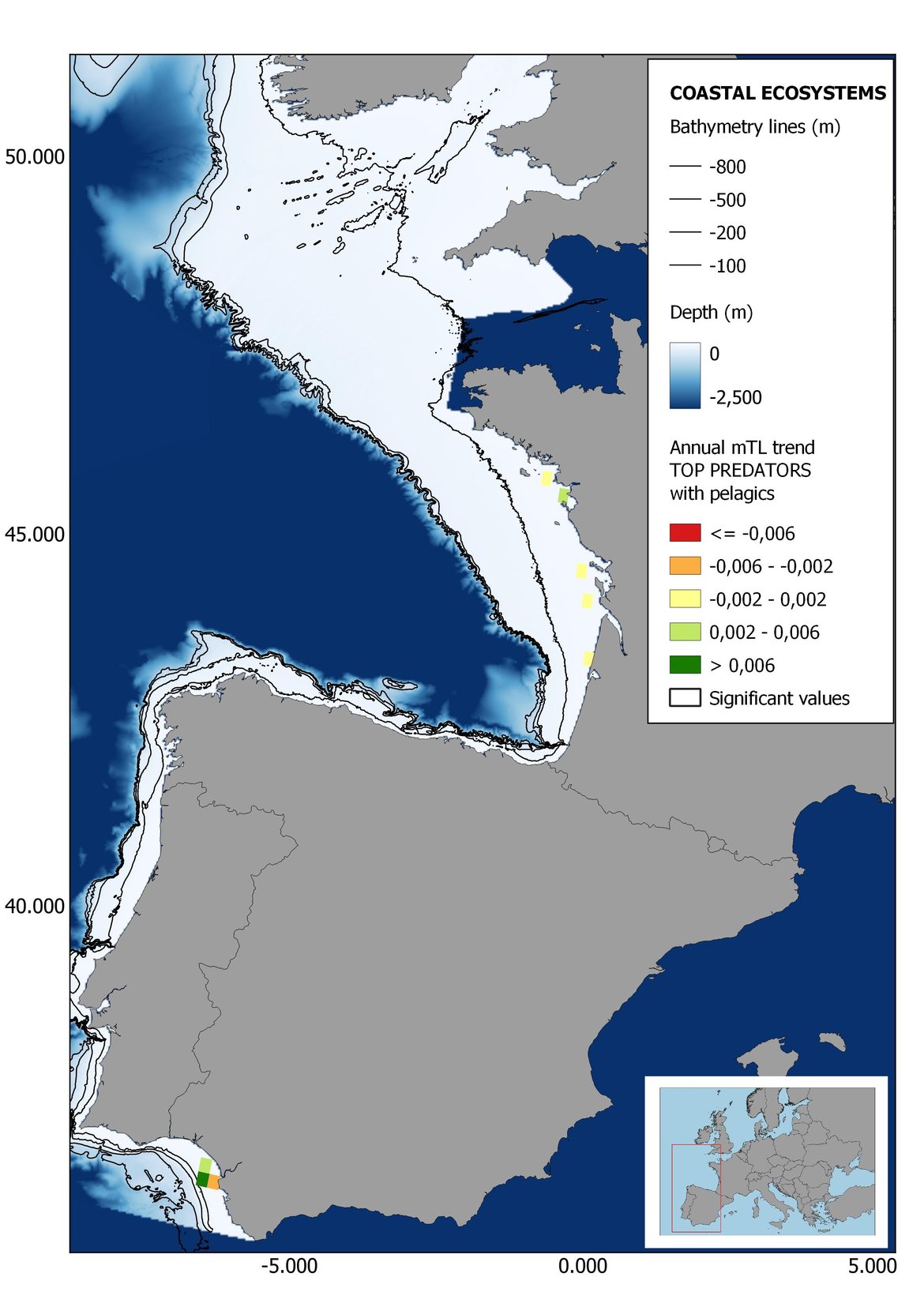
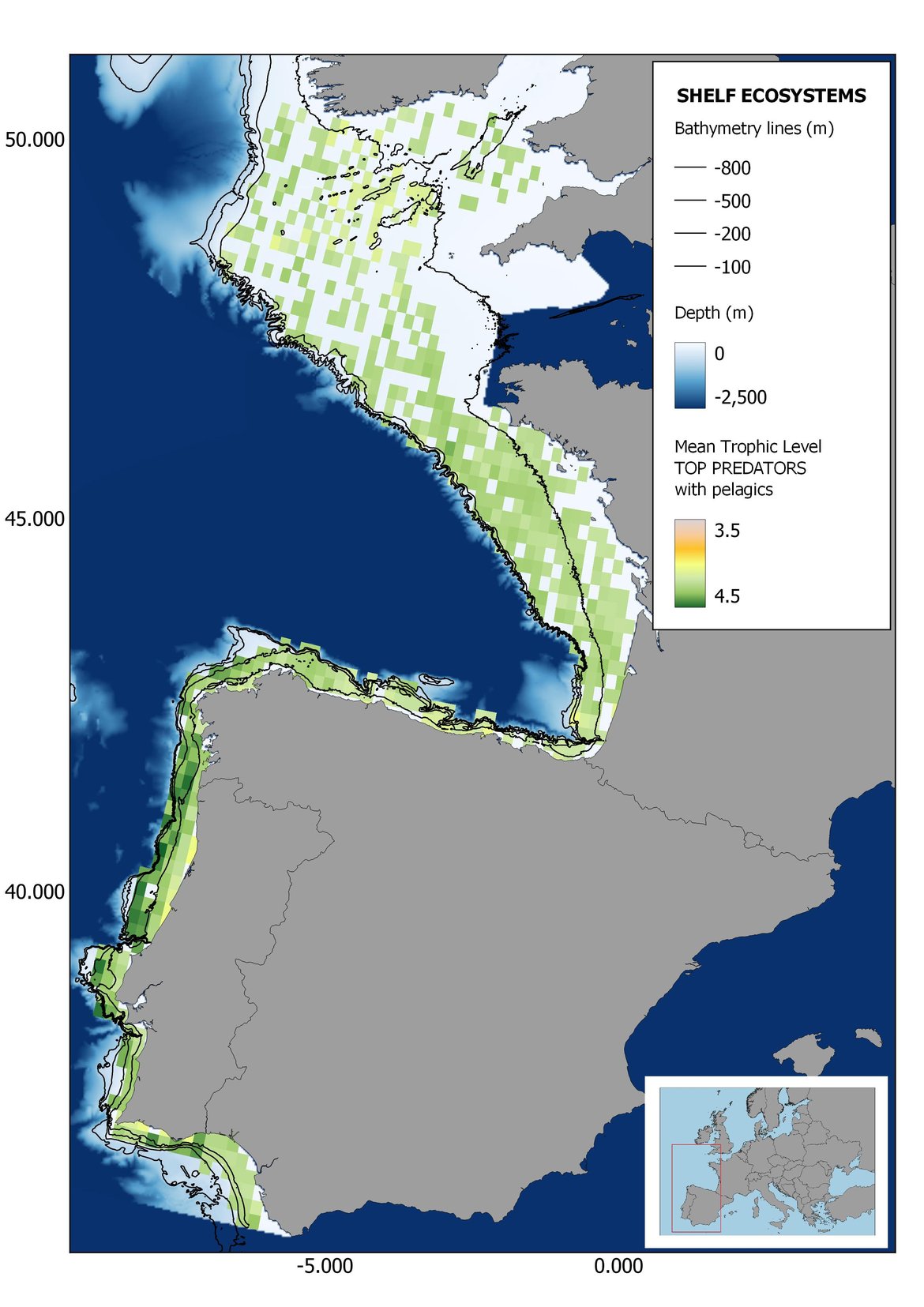
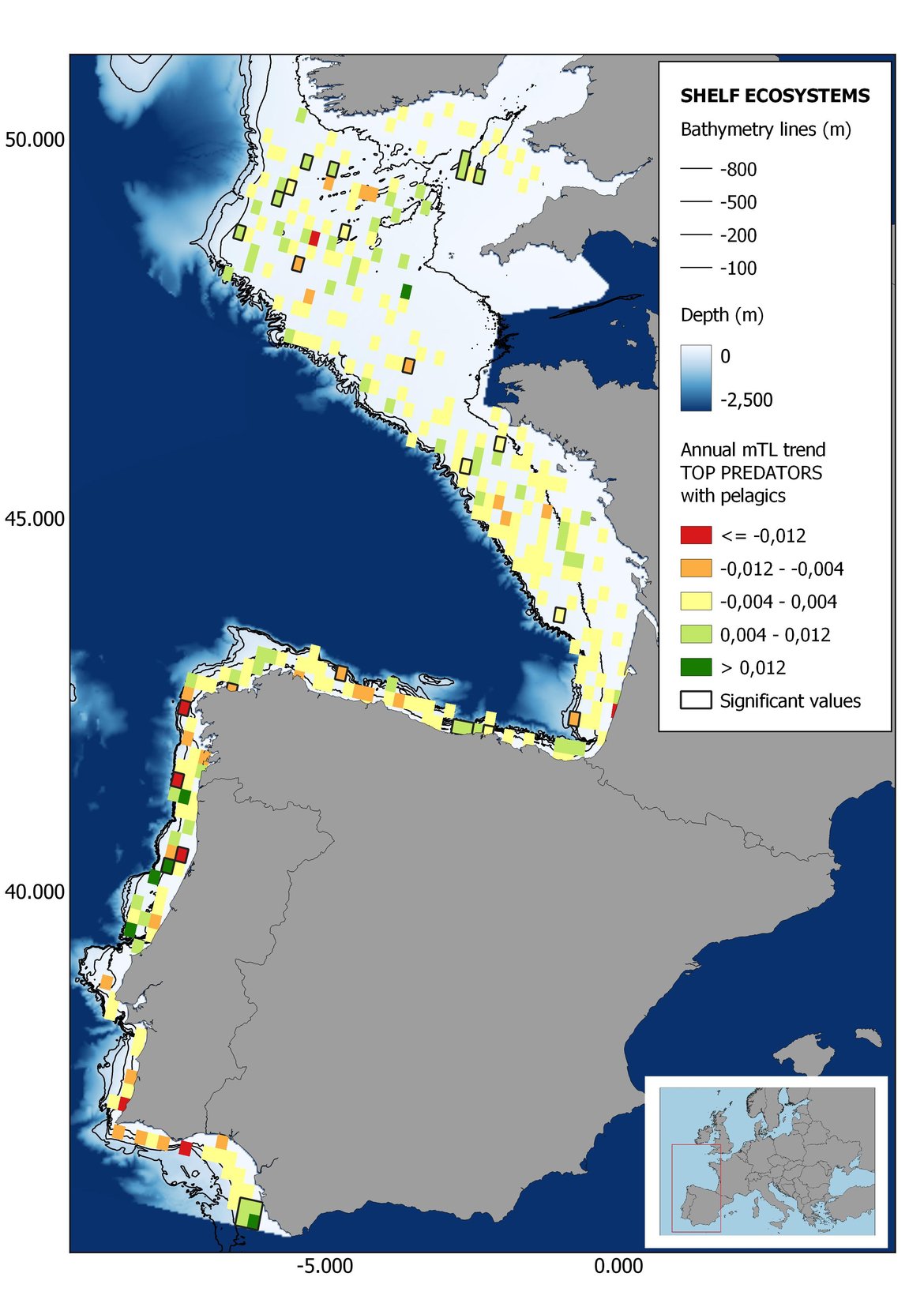
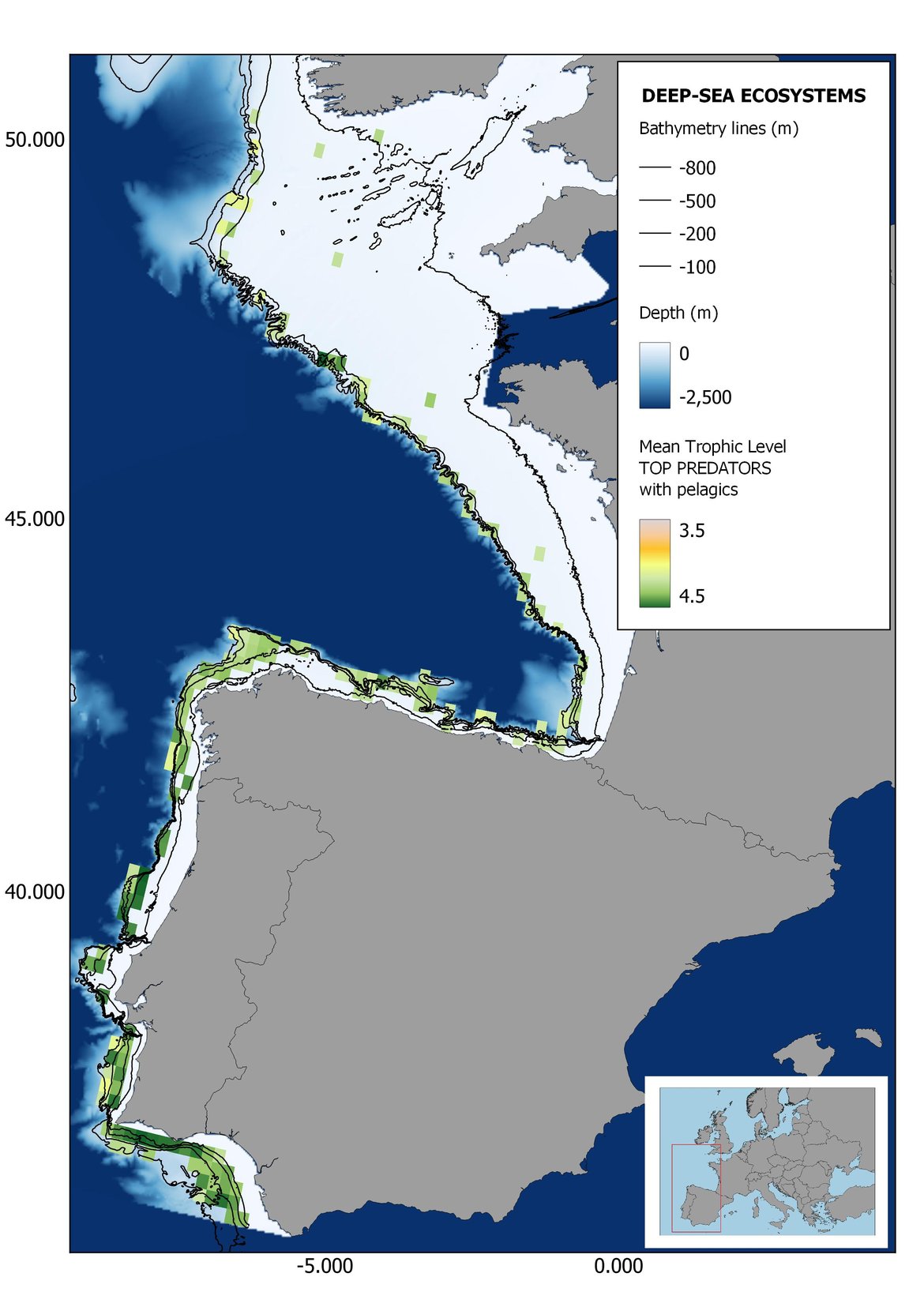
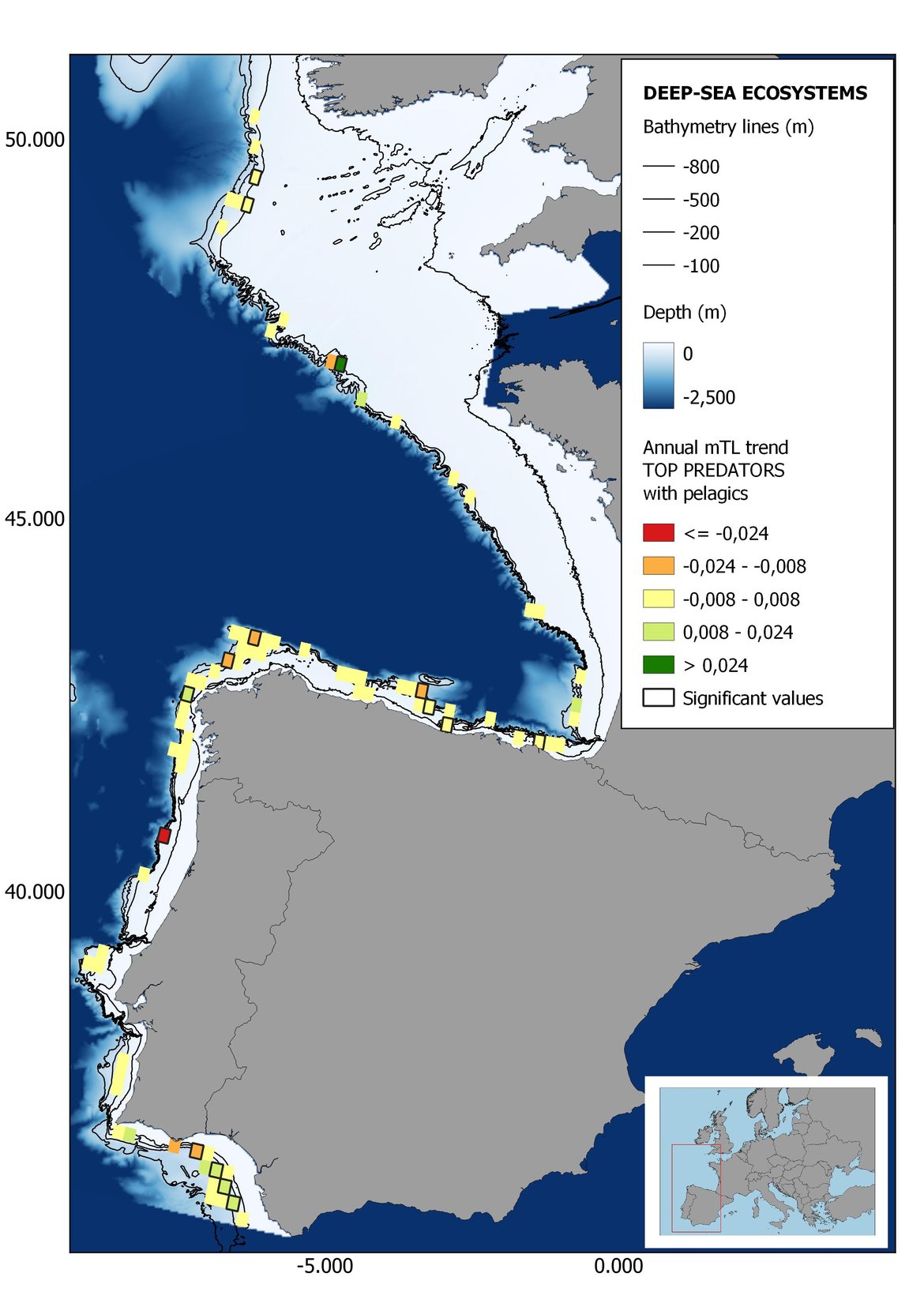
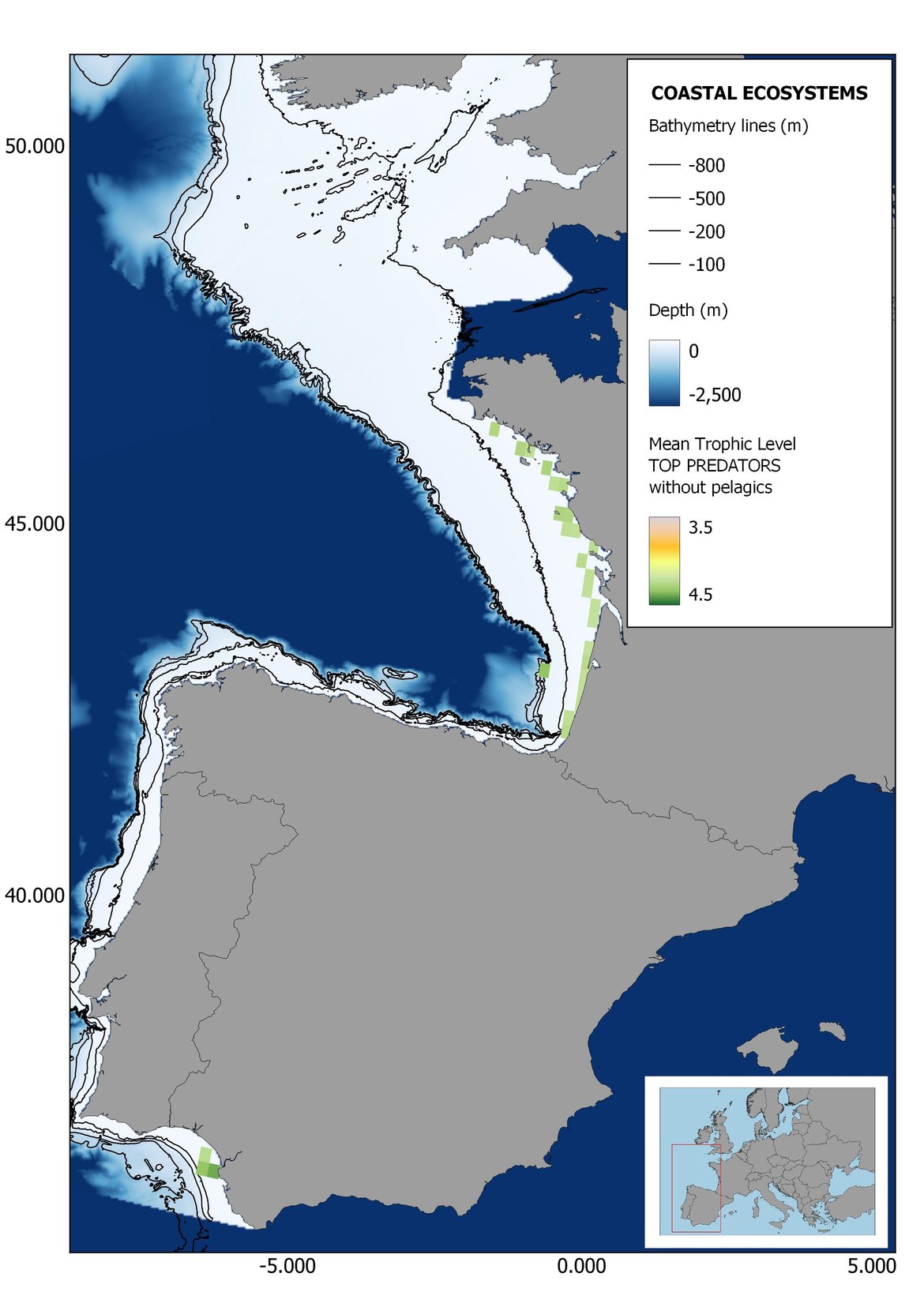
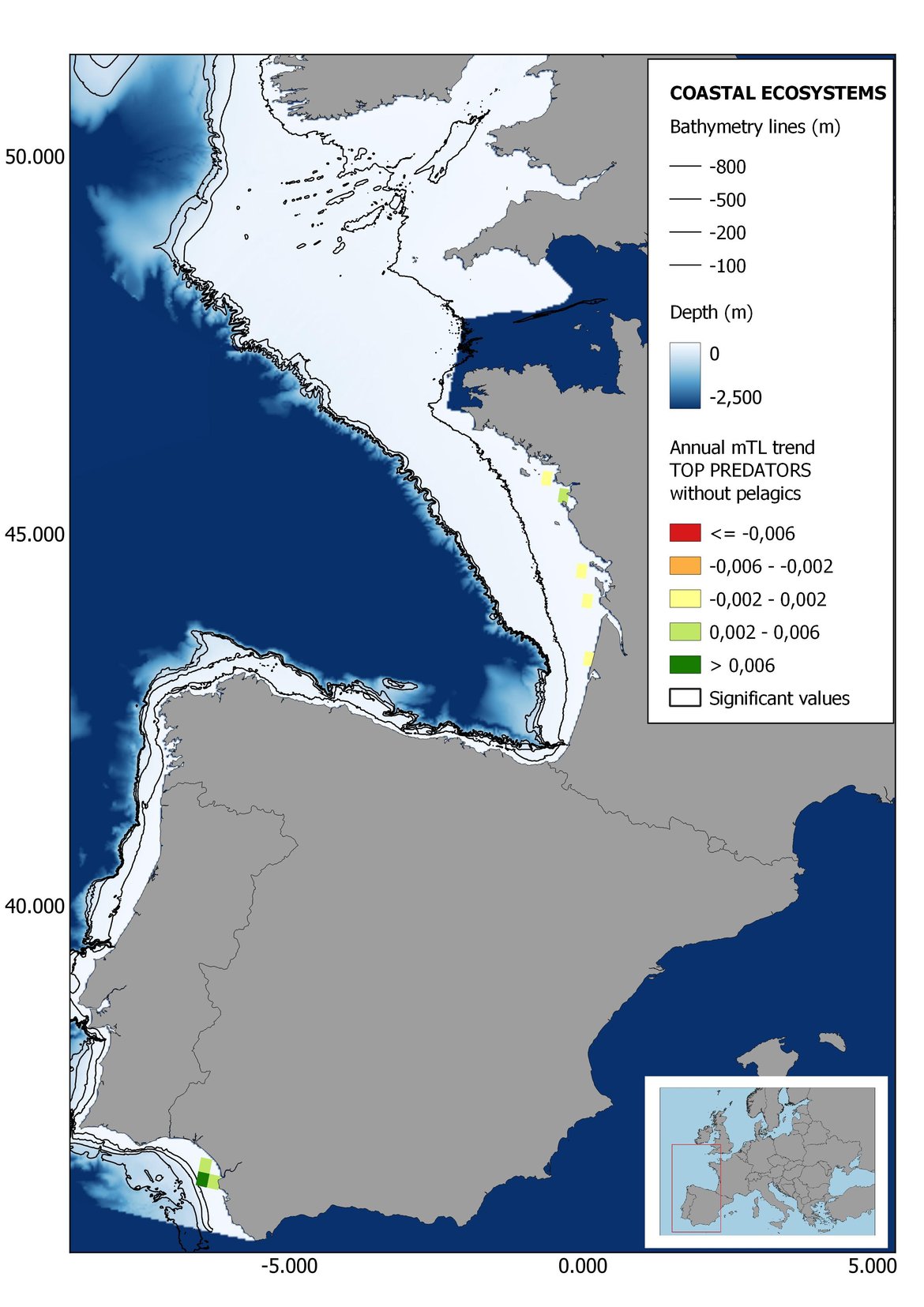
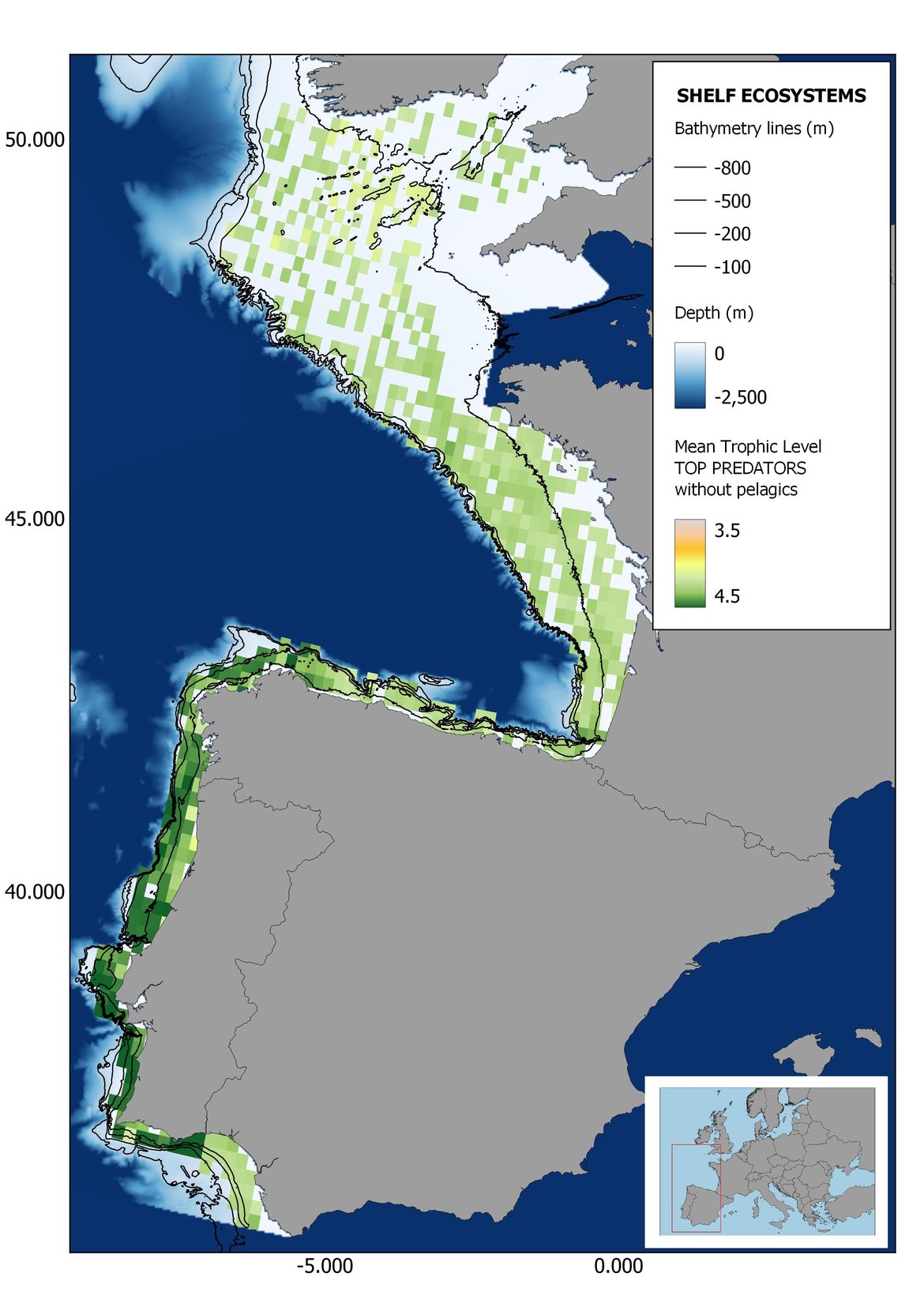
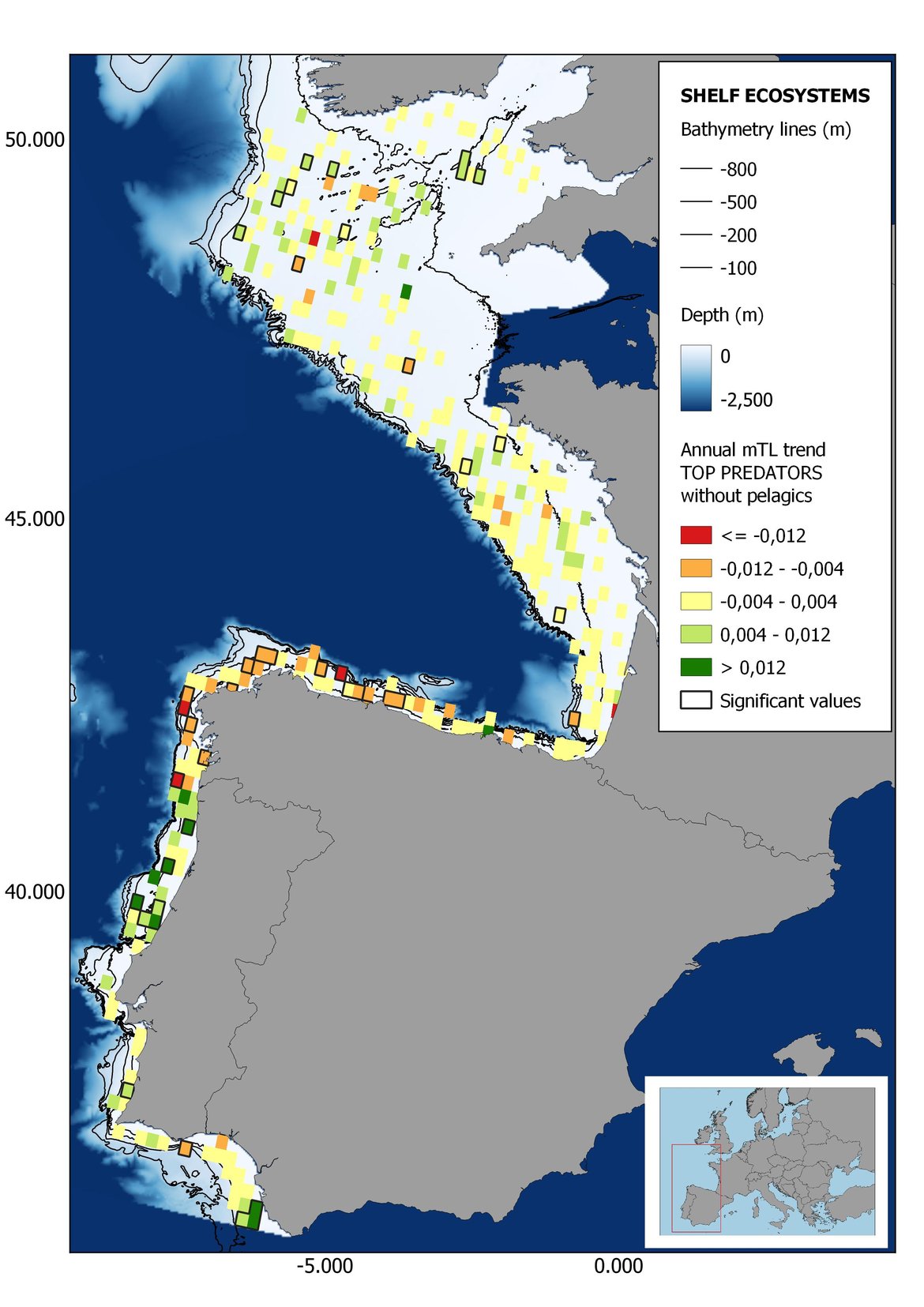
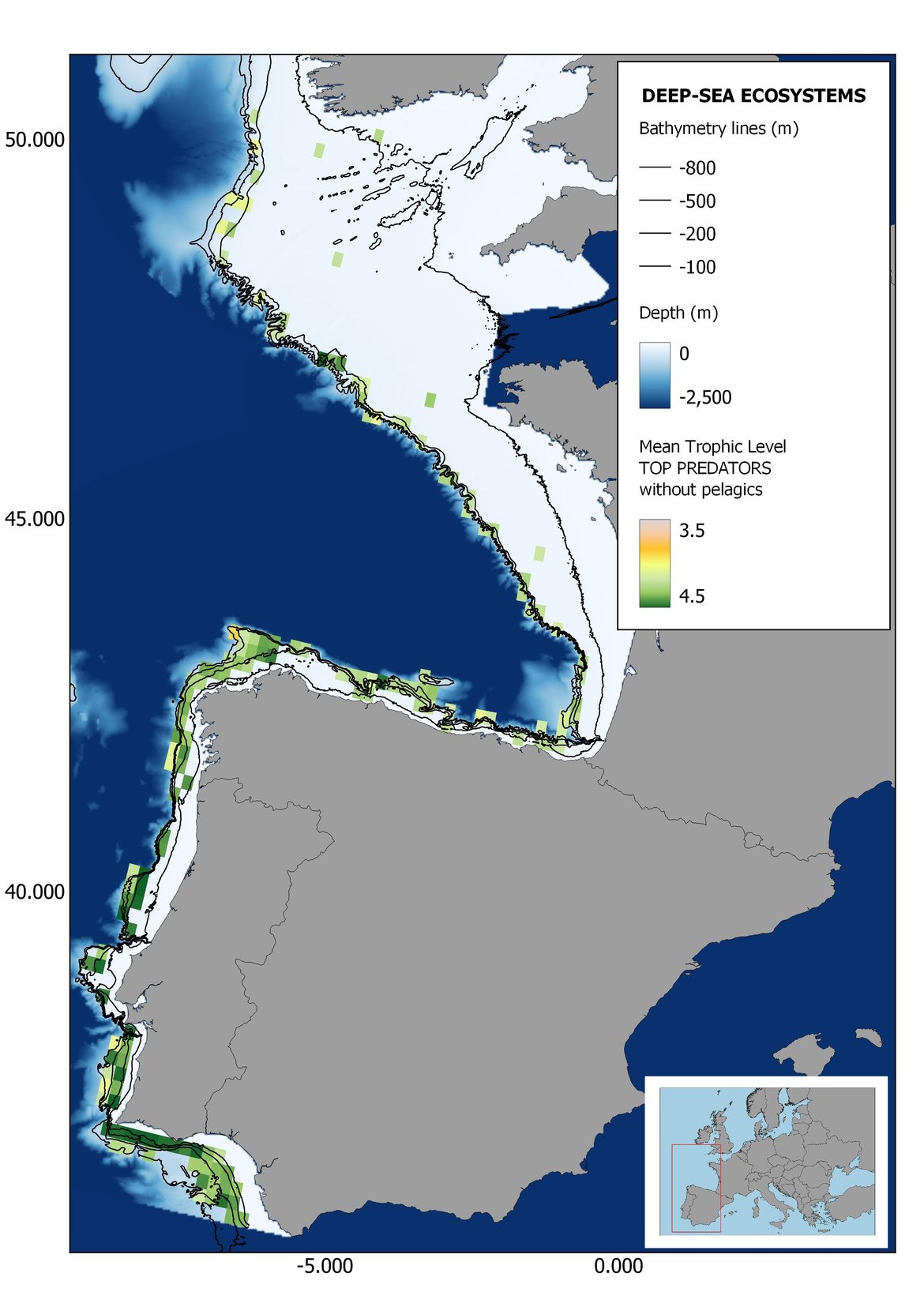
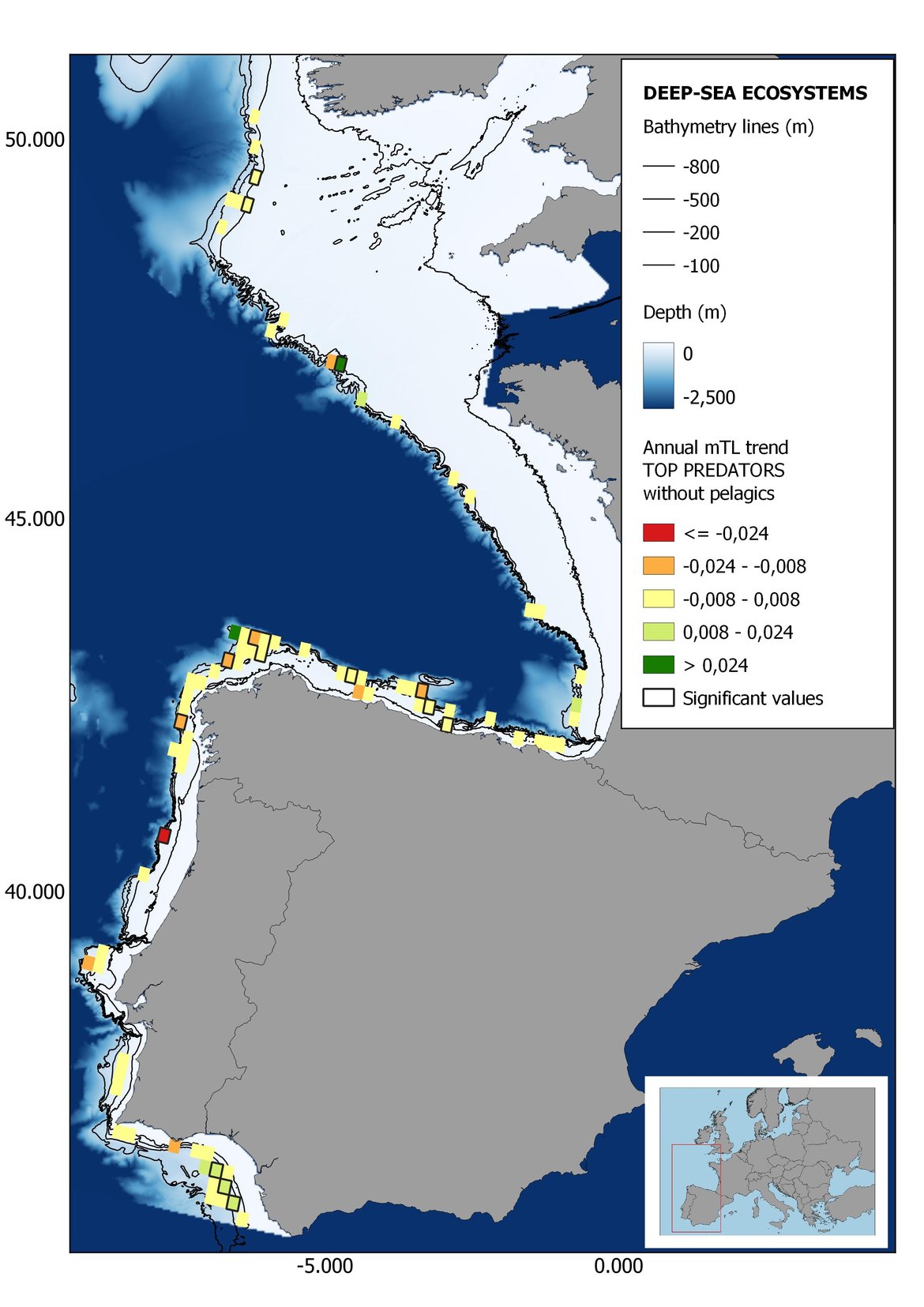
Figure 6: Maps showing the results of the spatio-temporal approach of Top predators: top 6 maps show mean Trophic Level by square, bottom 6 maps show trend of the MTL by square. Green = increasing trend, Orange-yellow = no trend, Red = decreasing trend. Significant trends are framed in bold.
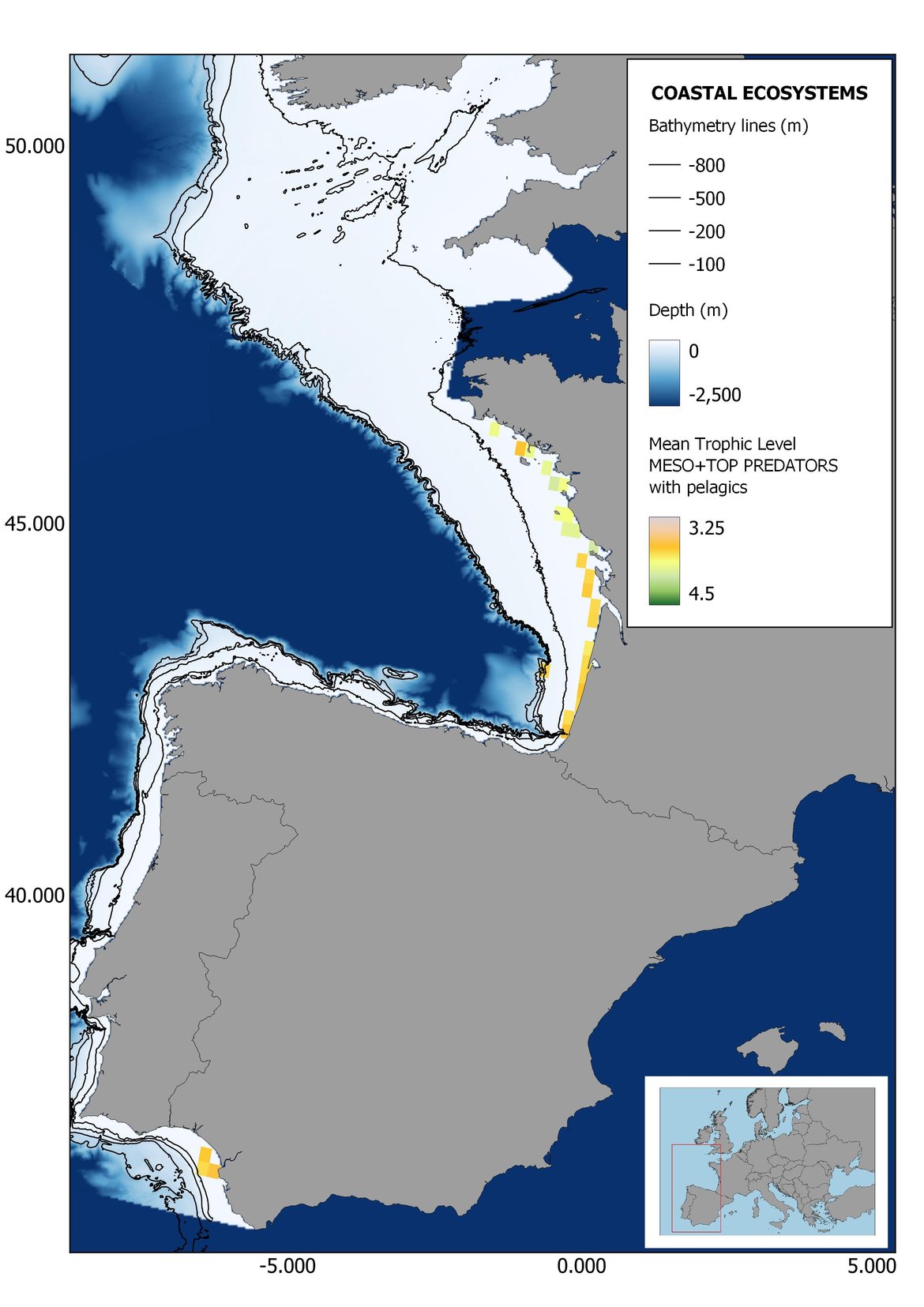
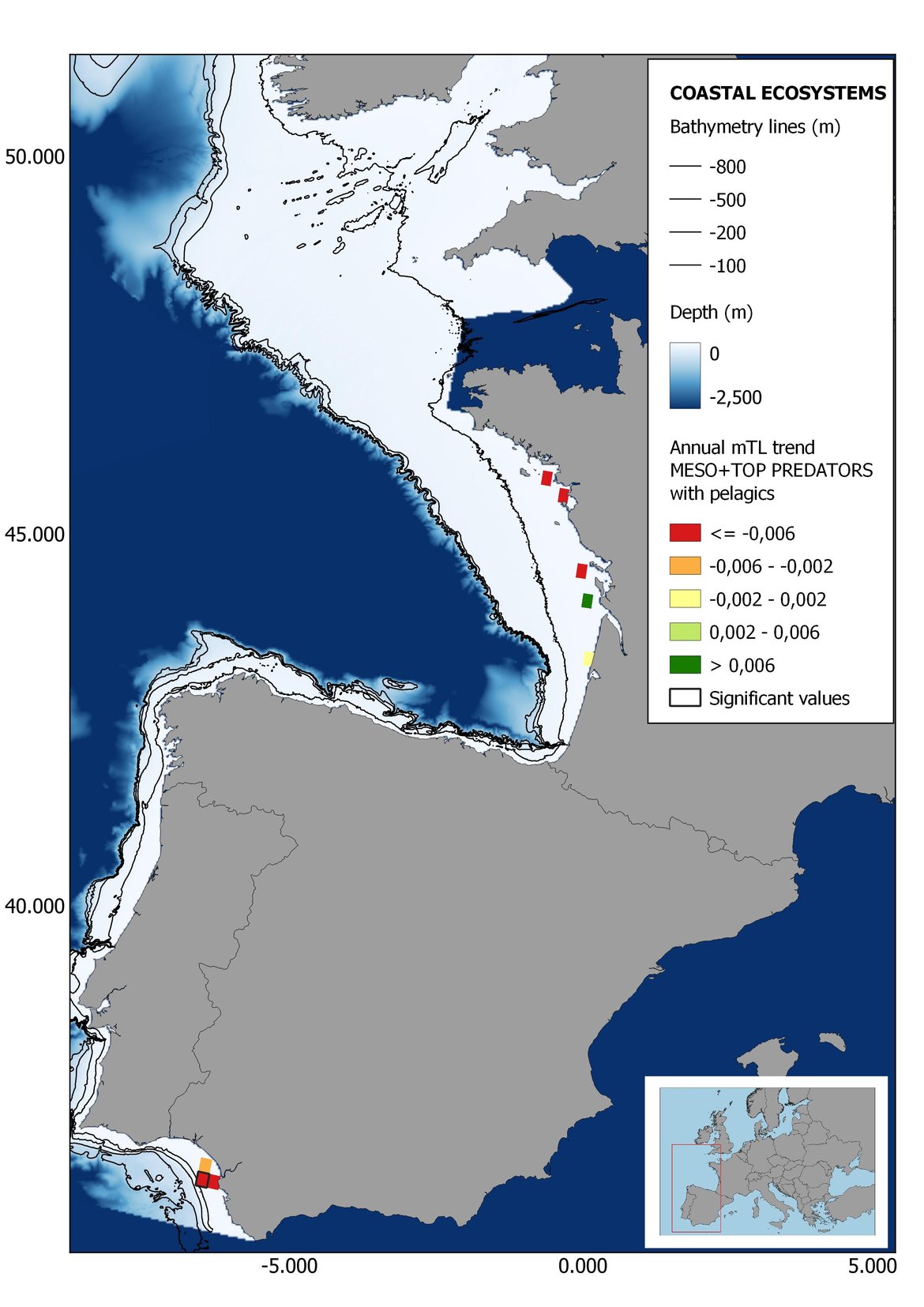
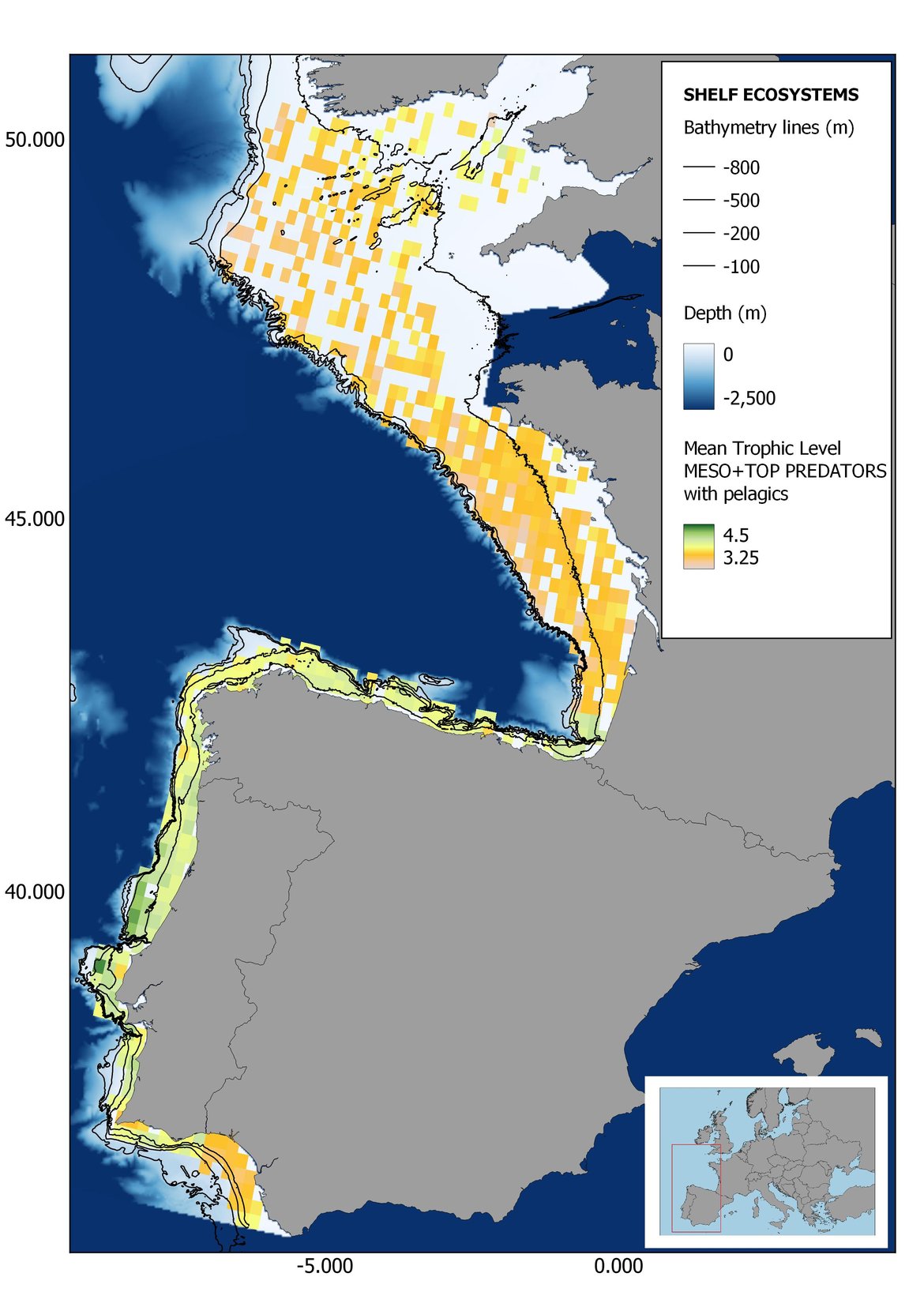
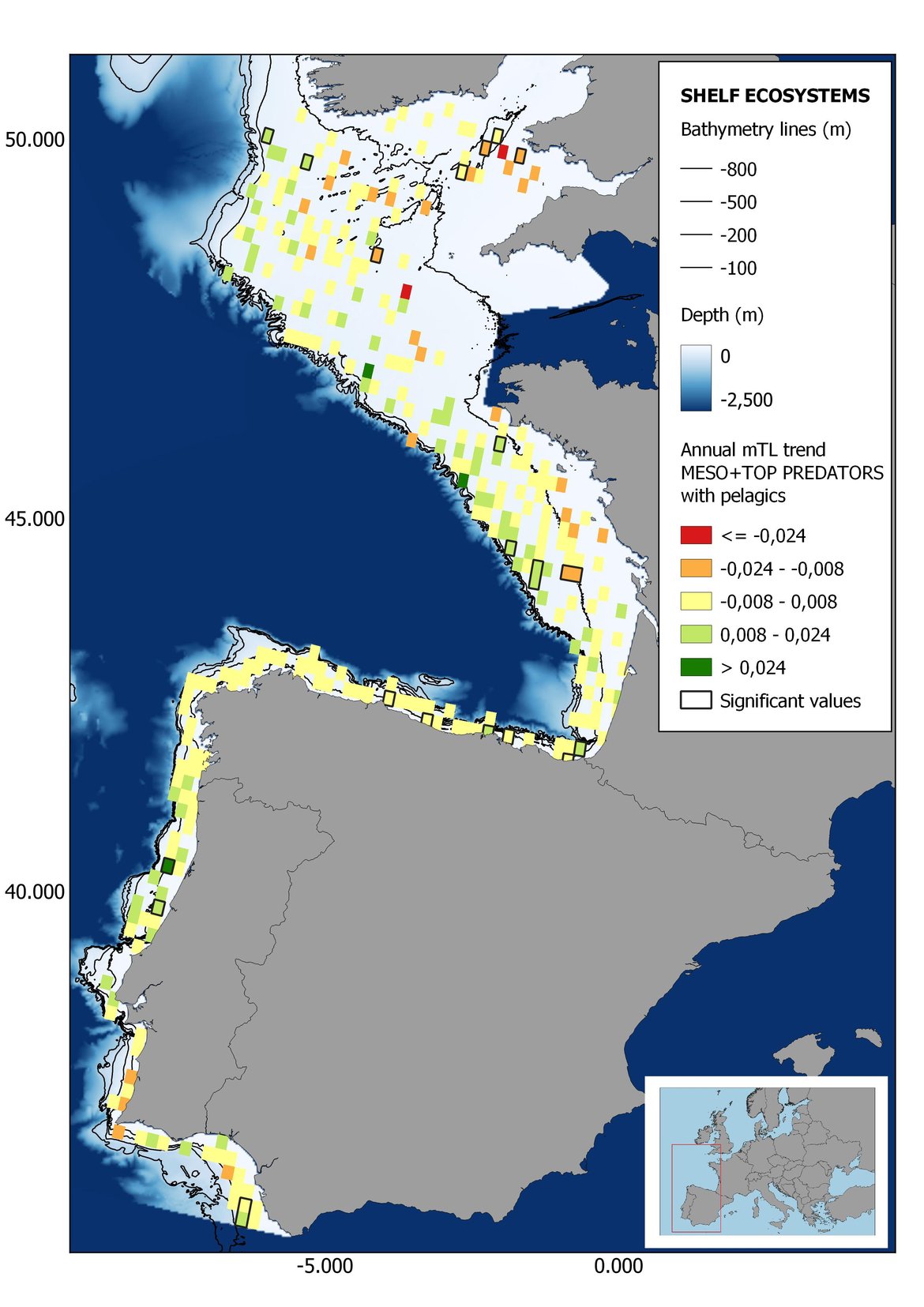
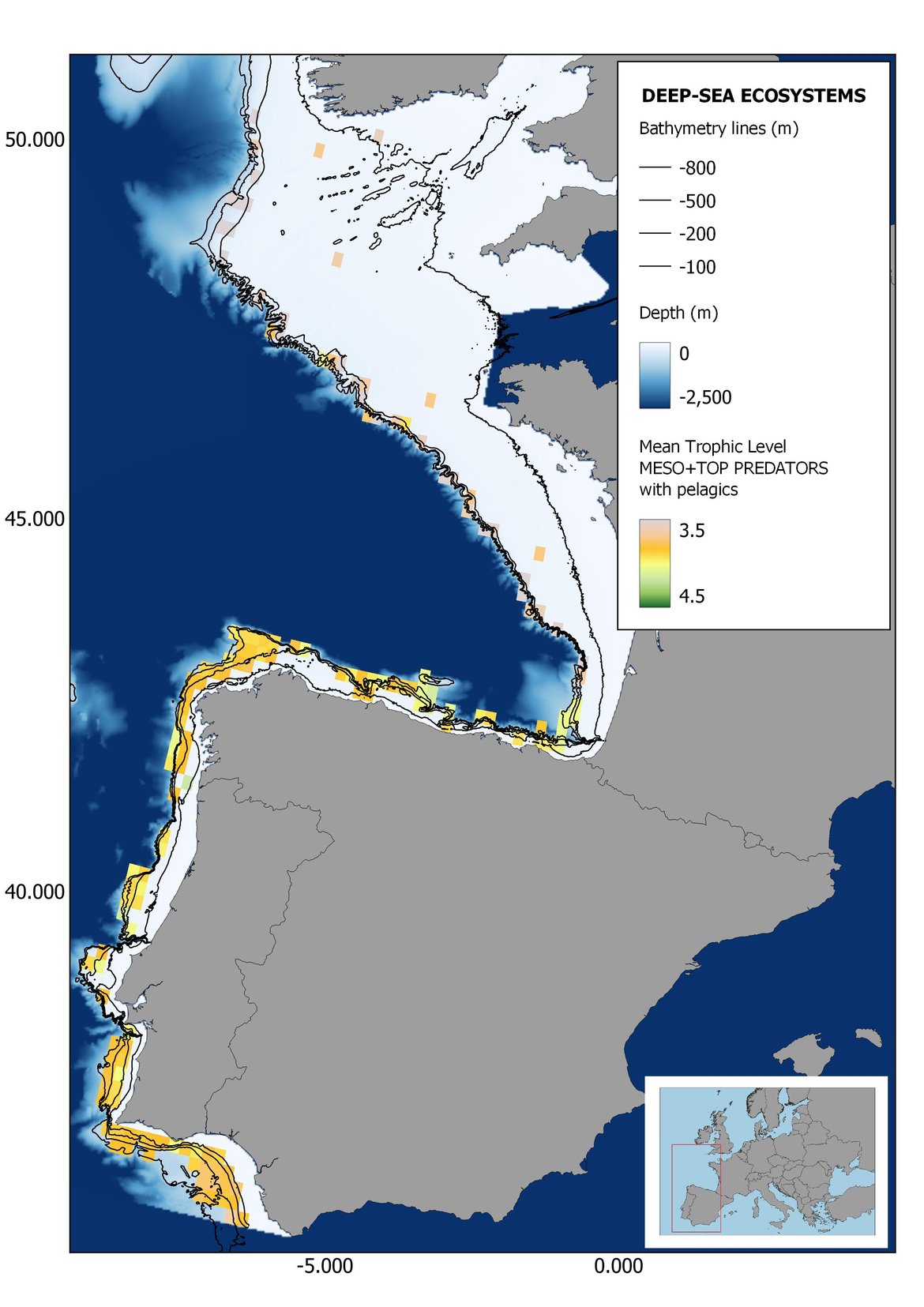
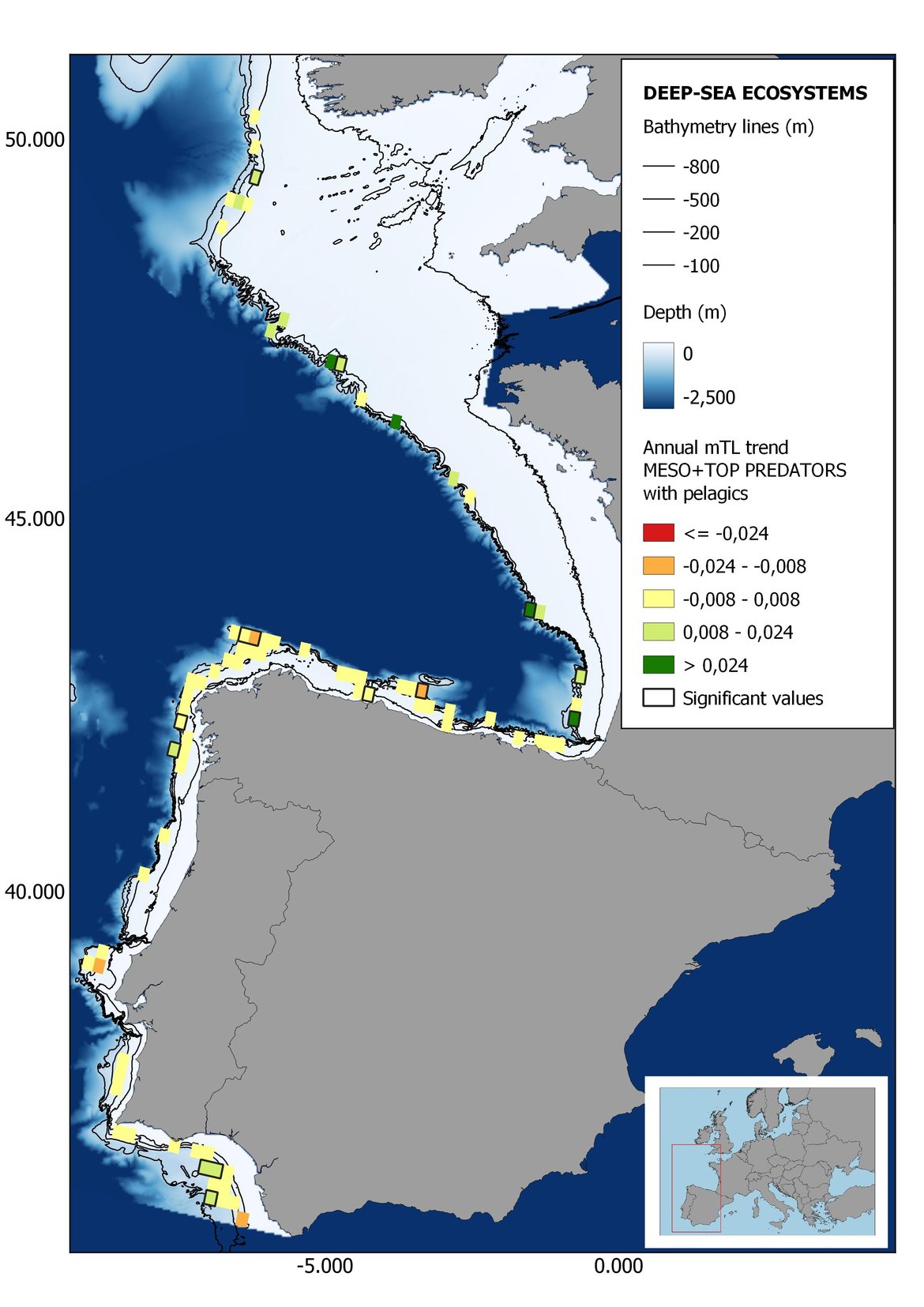
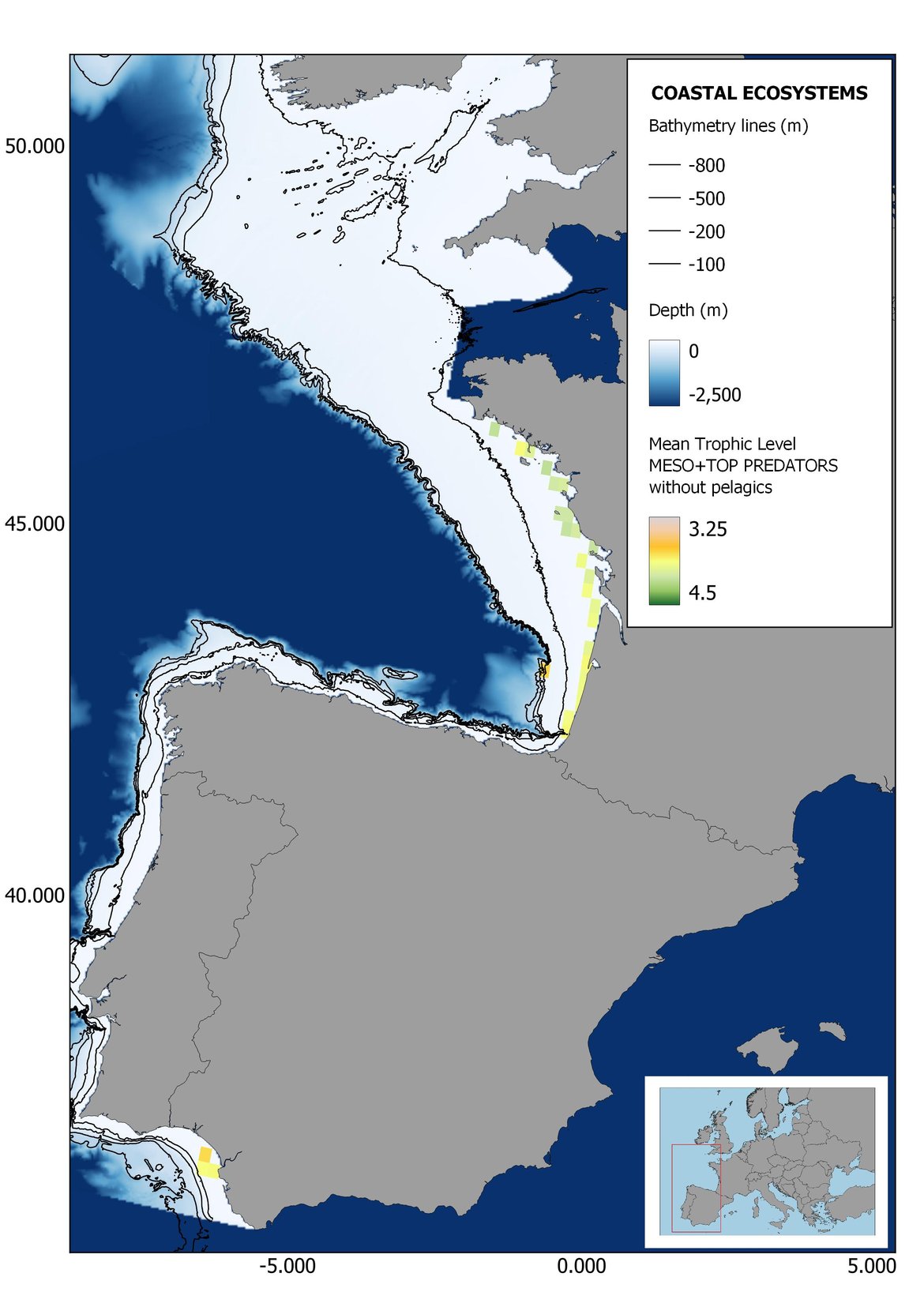
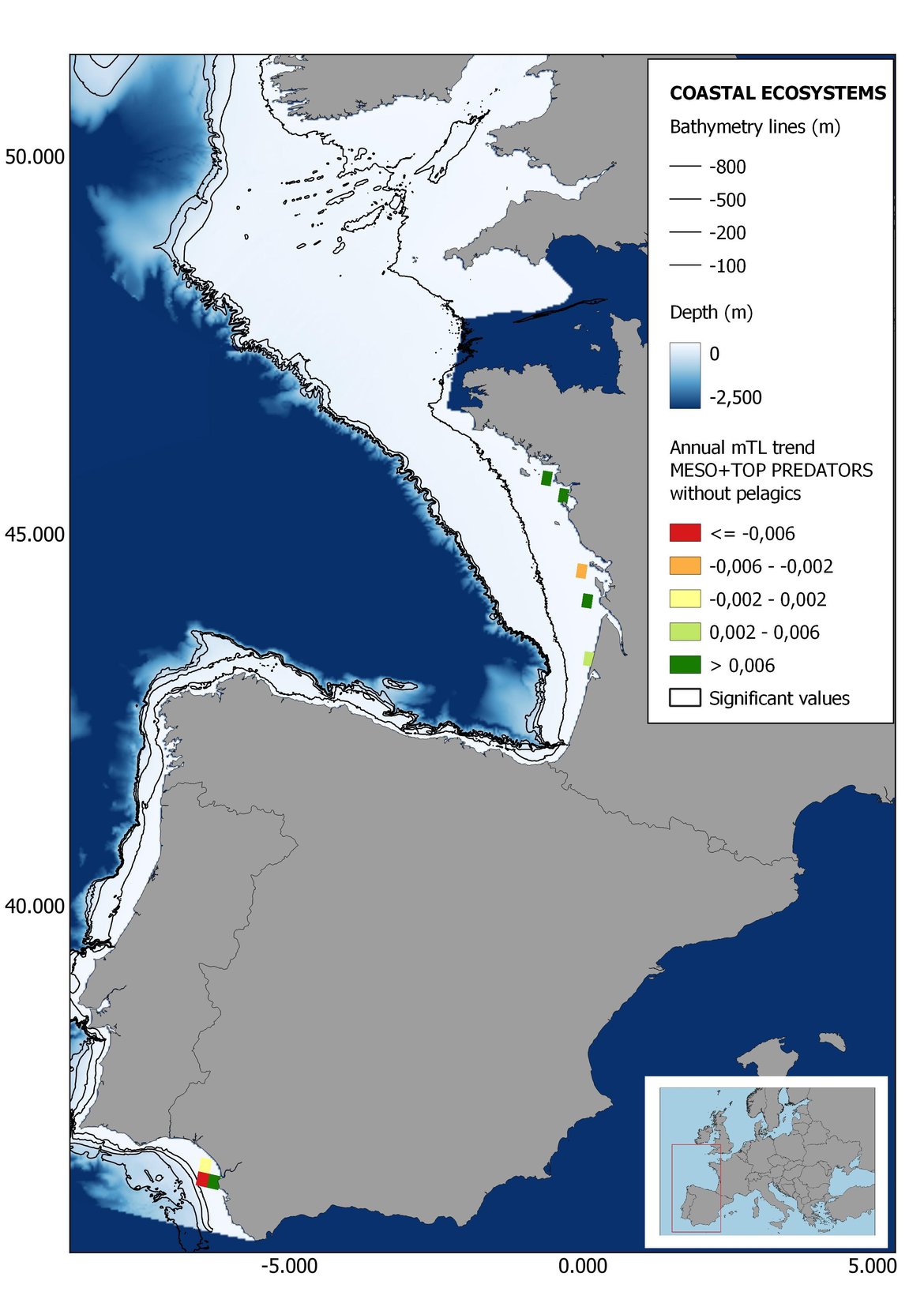
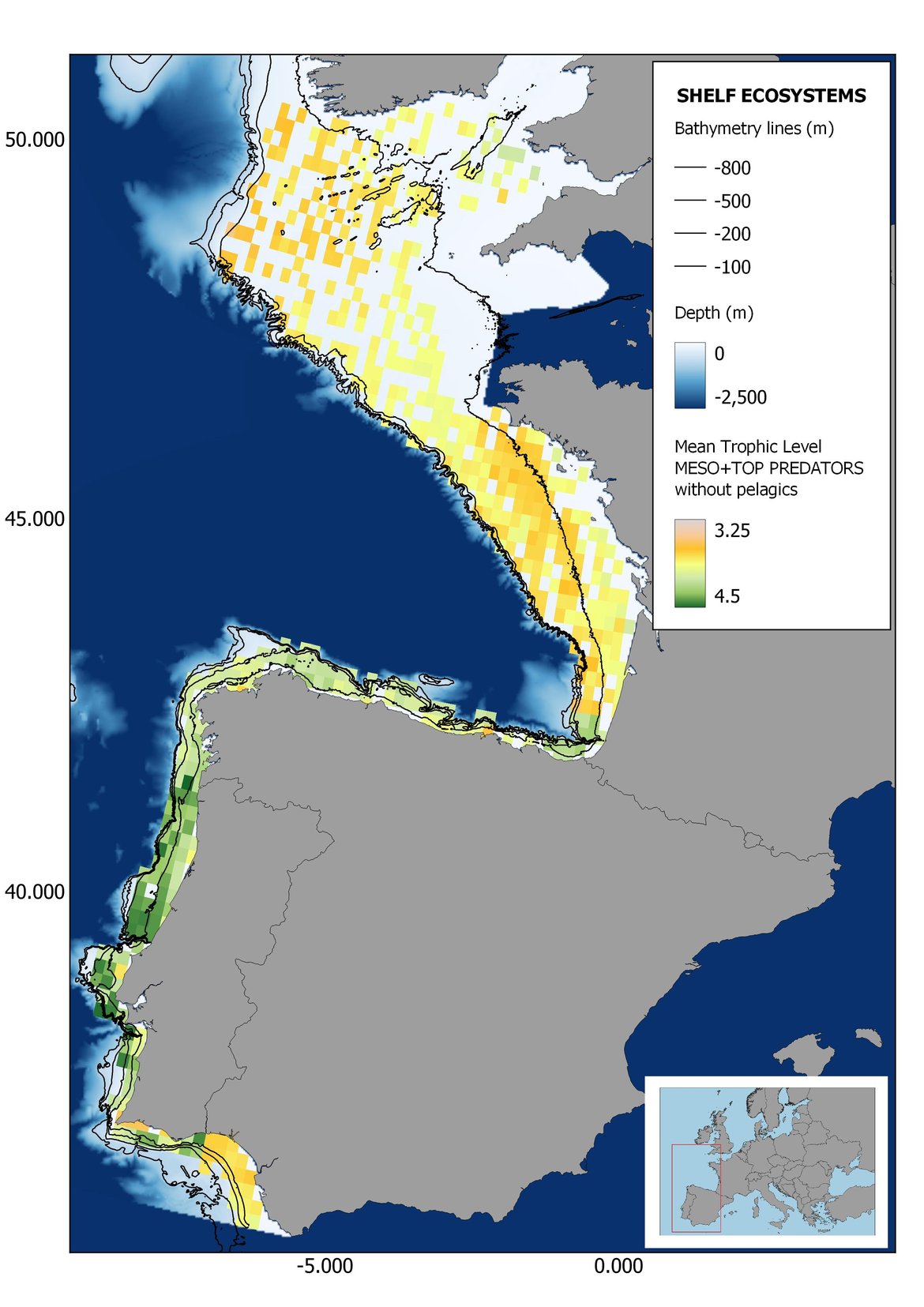
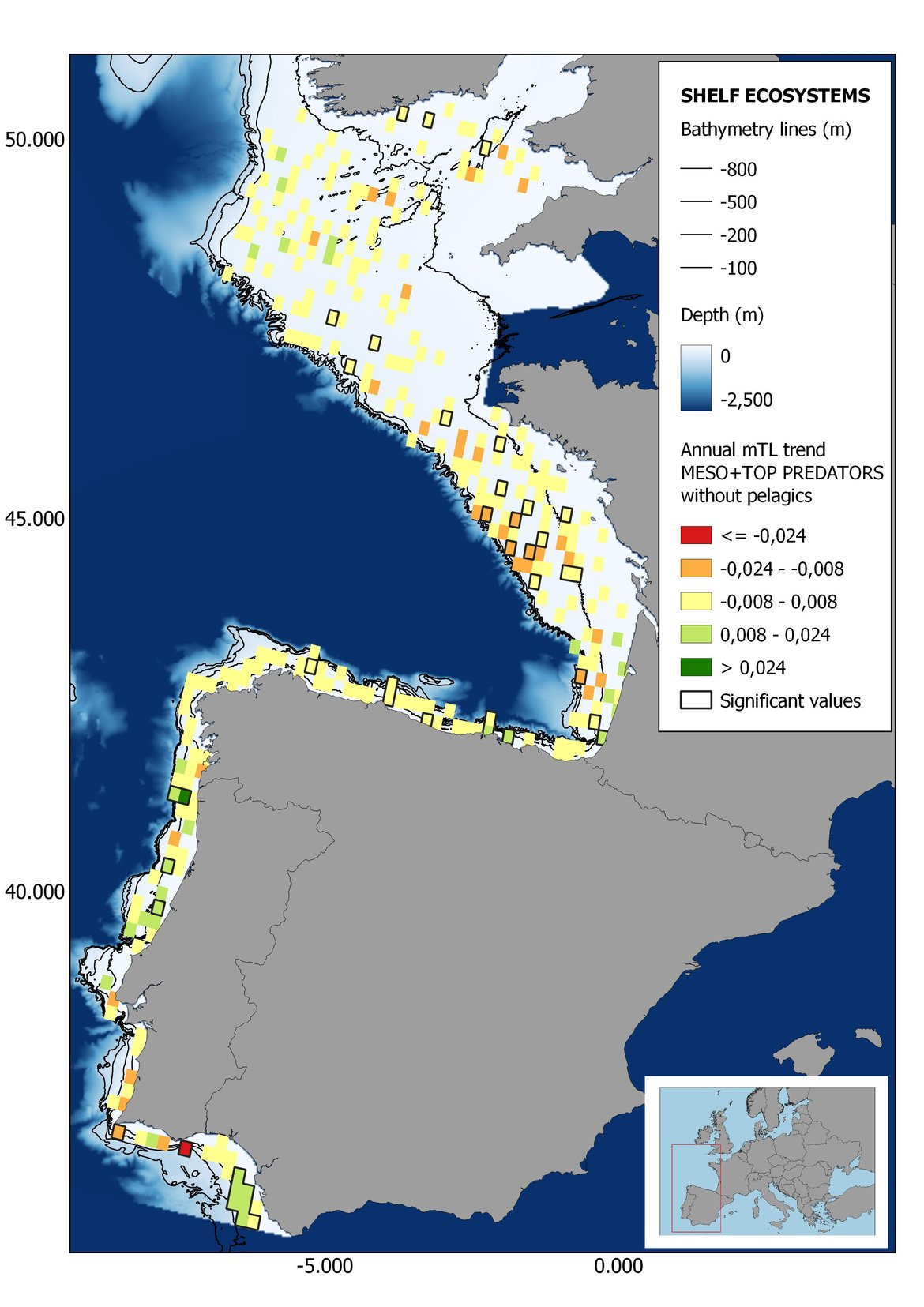
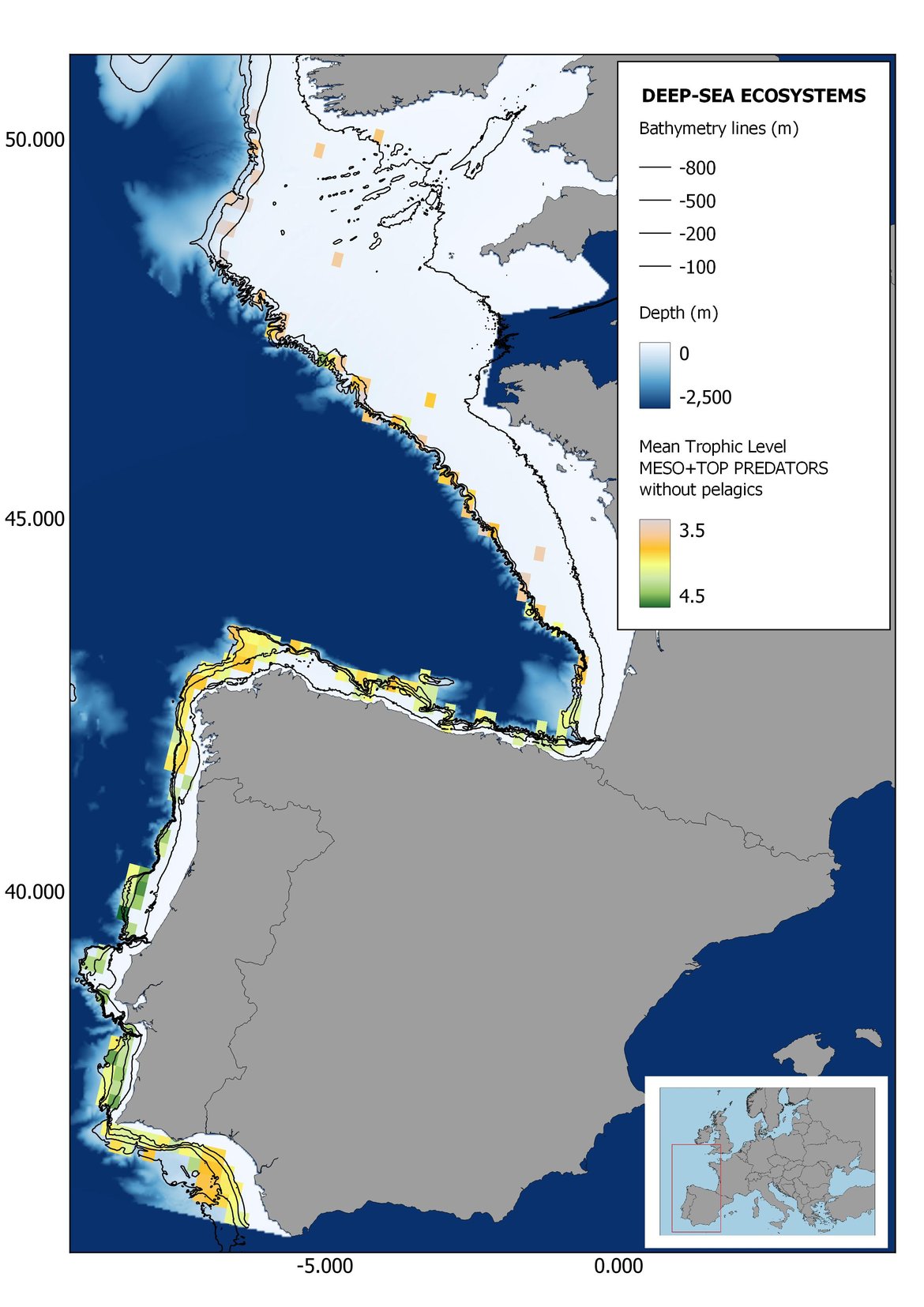
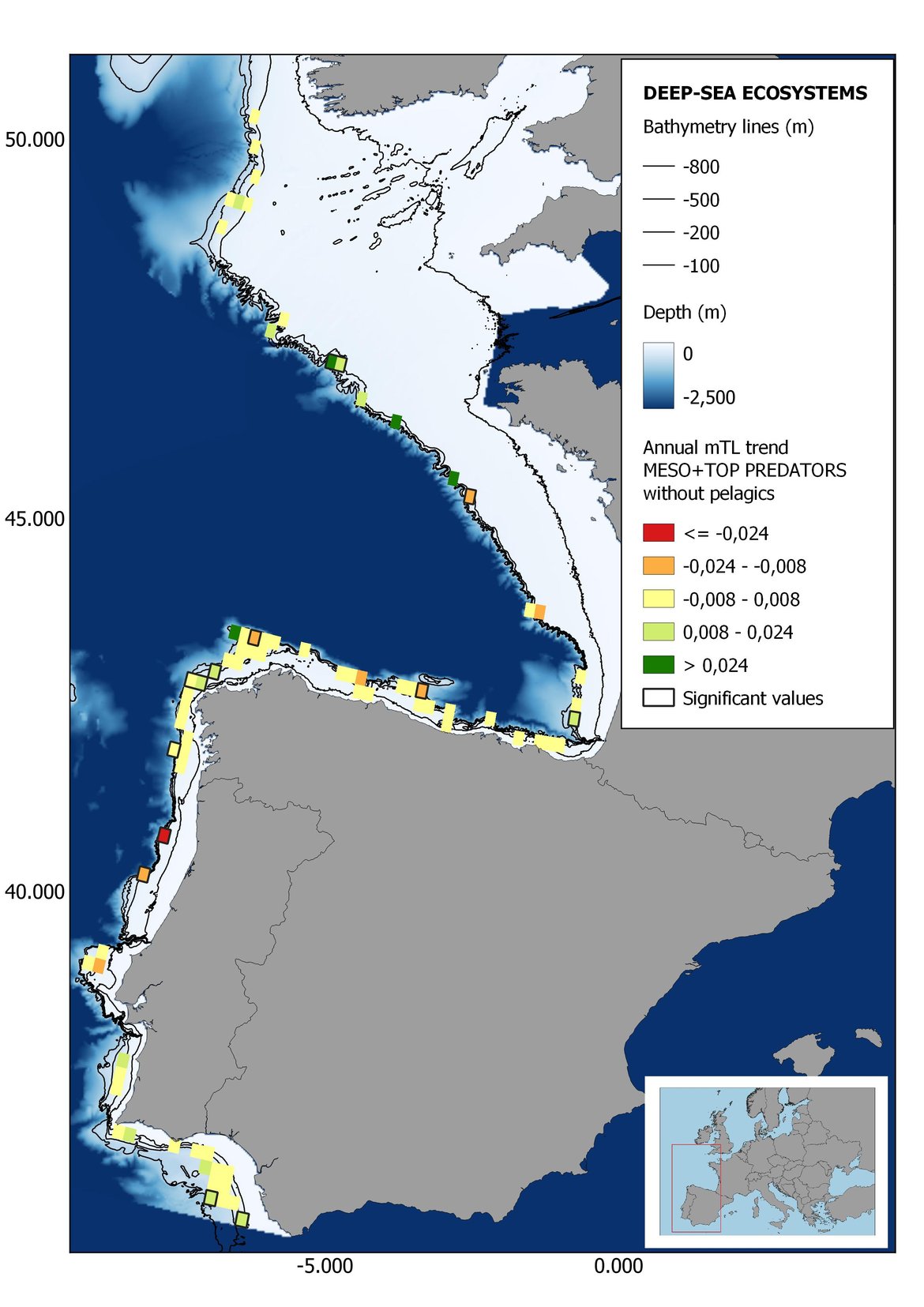
Figure 7: Maps showing the results of the spatio-temporal approach of Meso + Top predators: top 6 maps show mean Trophic Level by square, bottom 6 maps show trend of the MTL by square. Green = increasing trend, Orange-yellow = no trend, Red = decreasing trend. Significant trends are framed in bold.
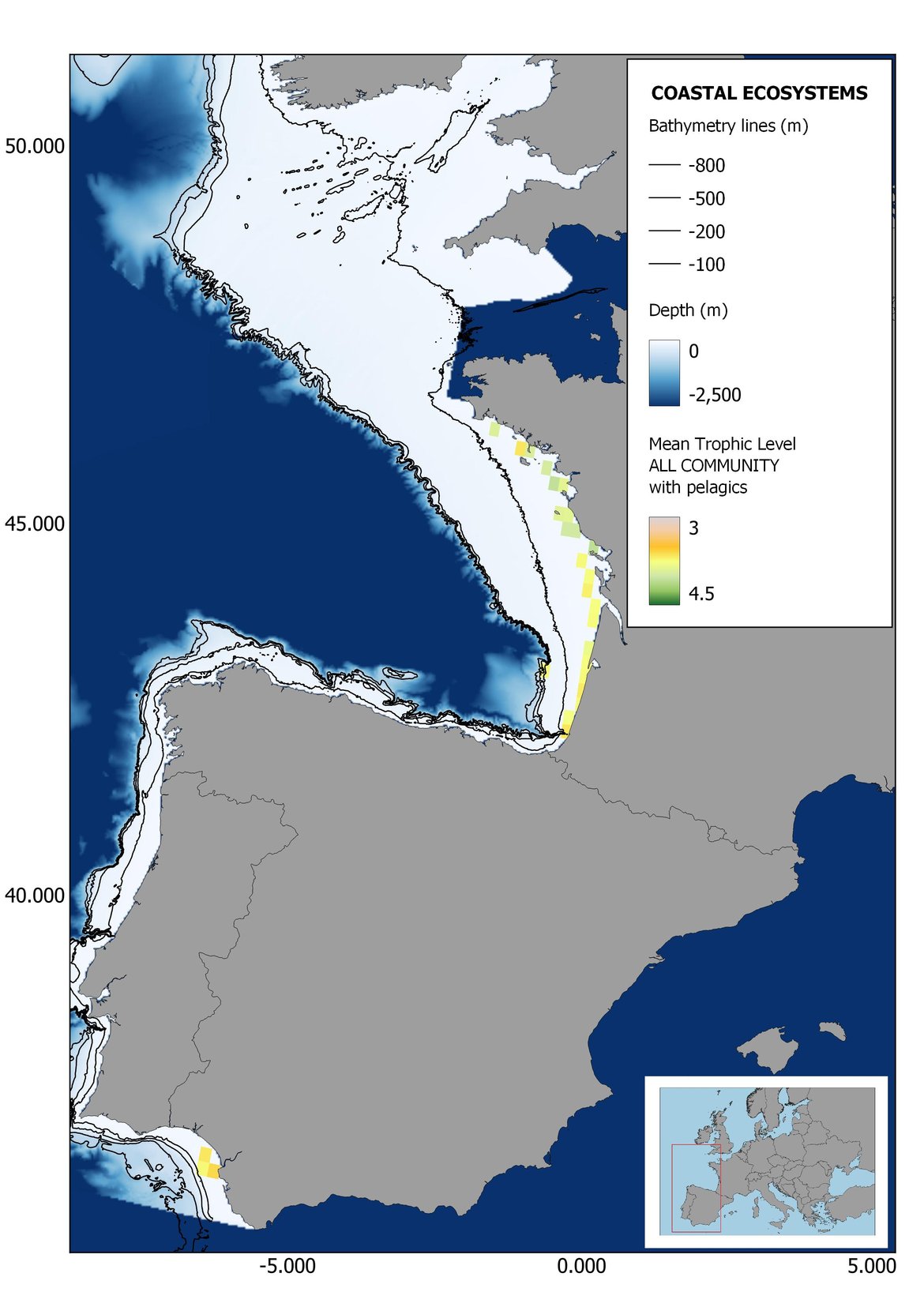
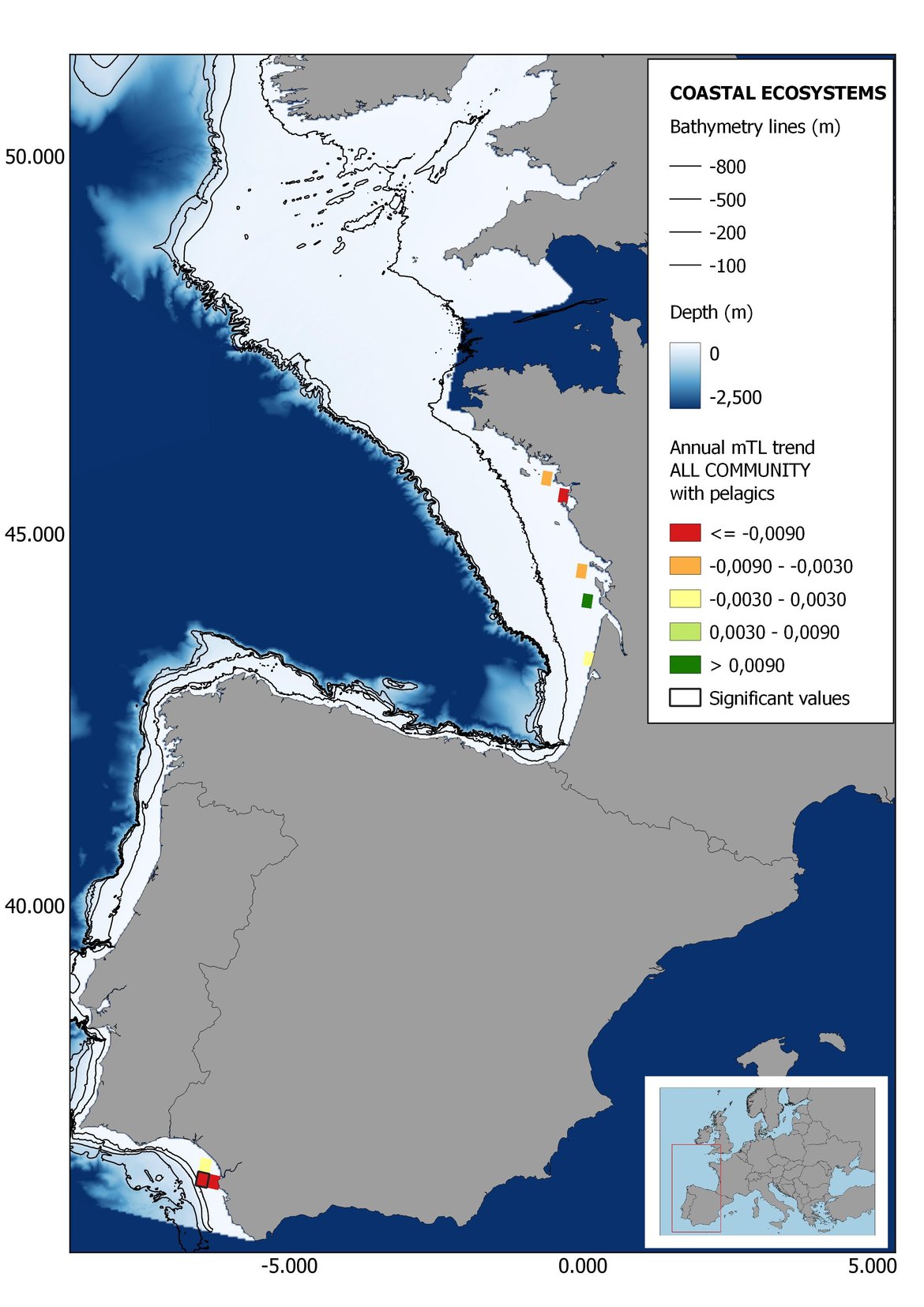
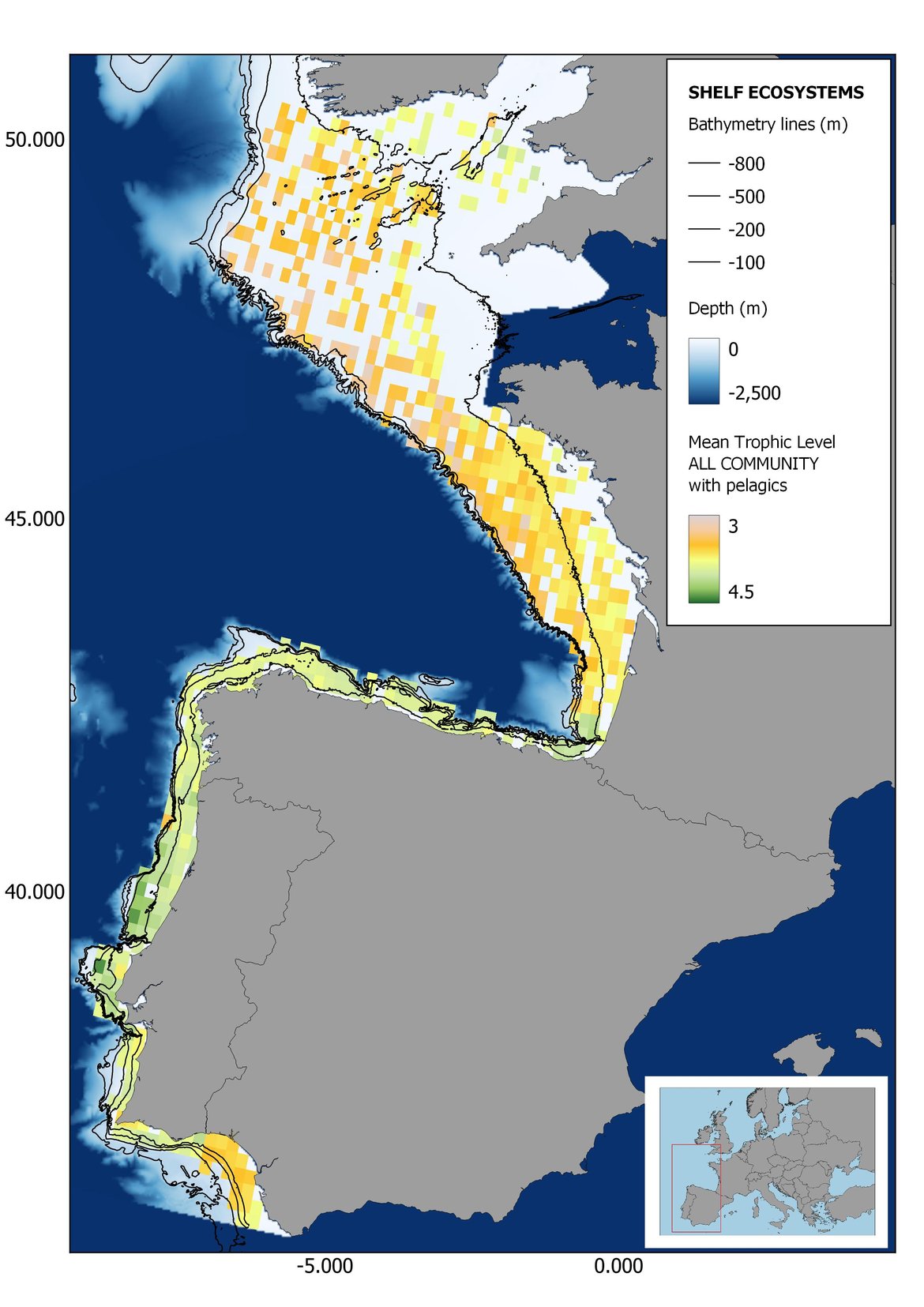
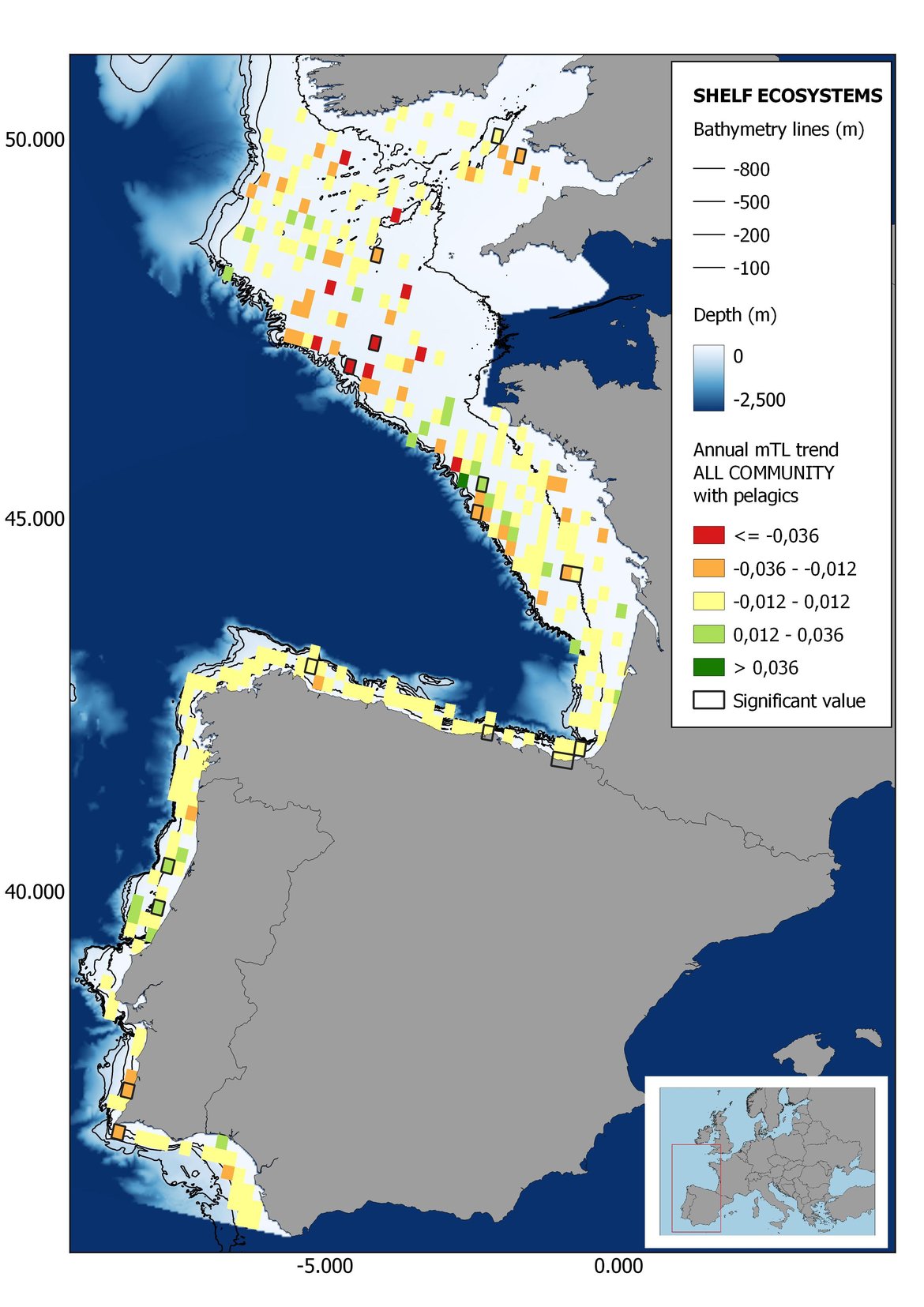
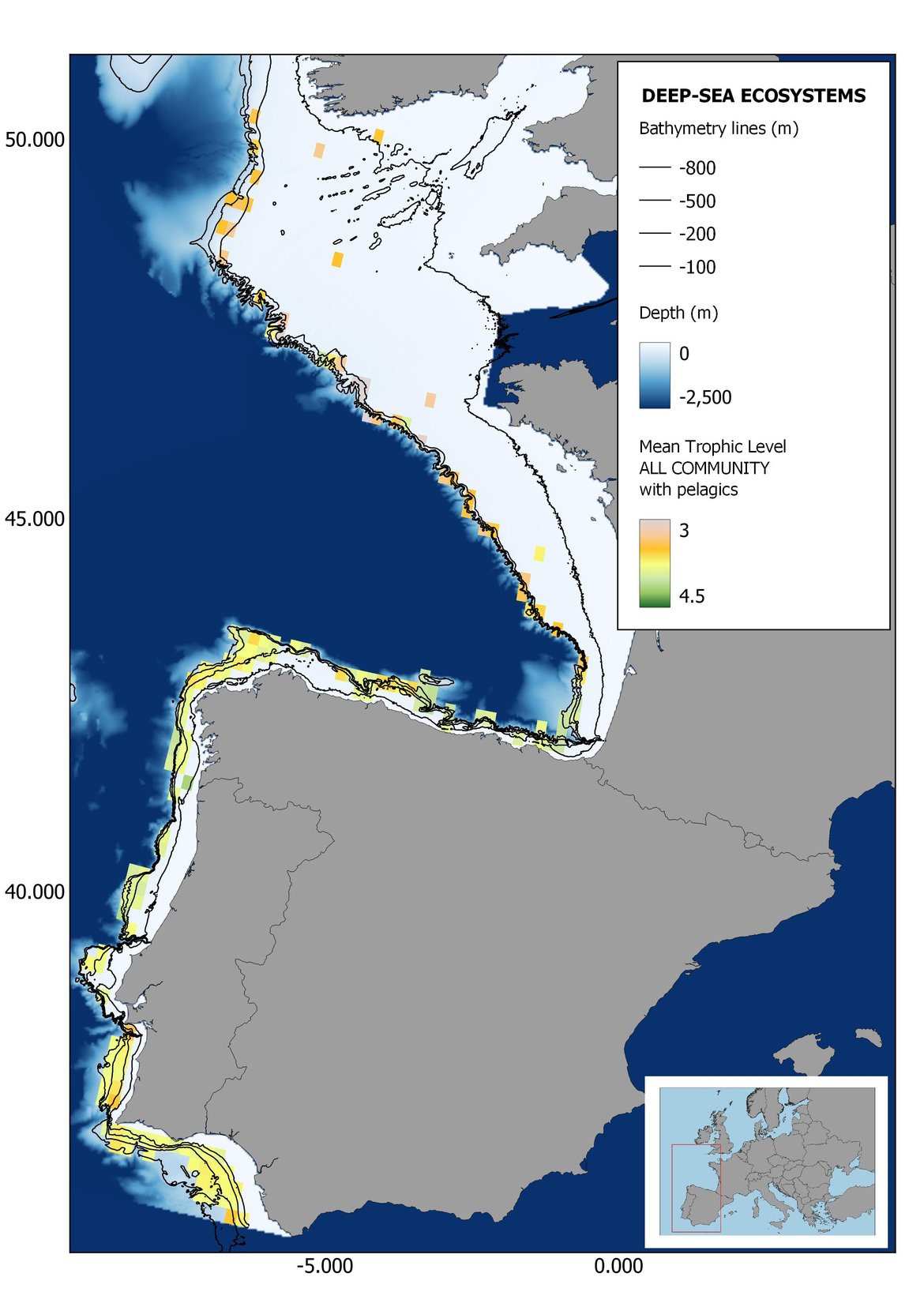
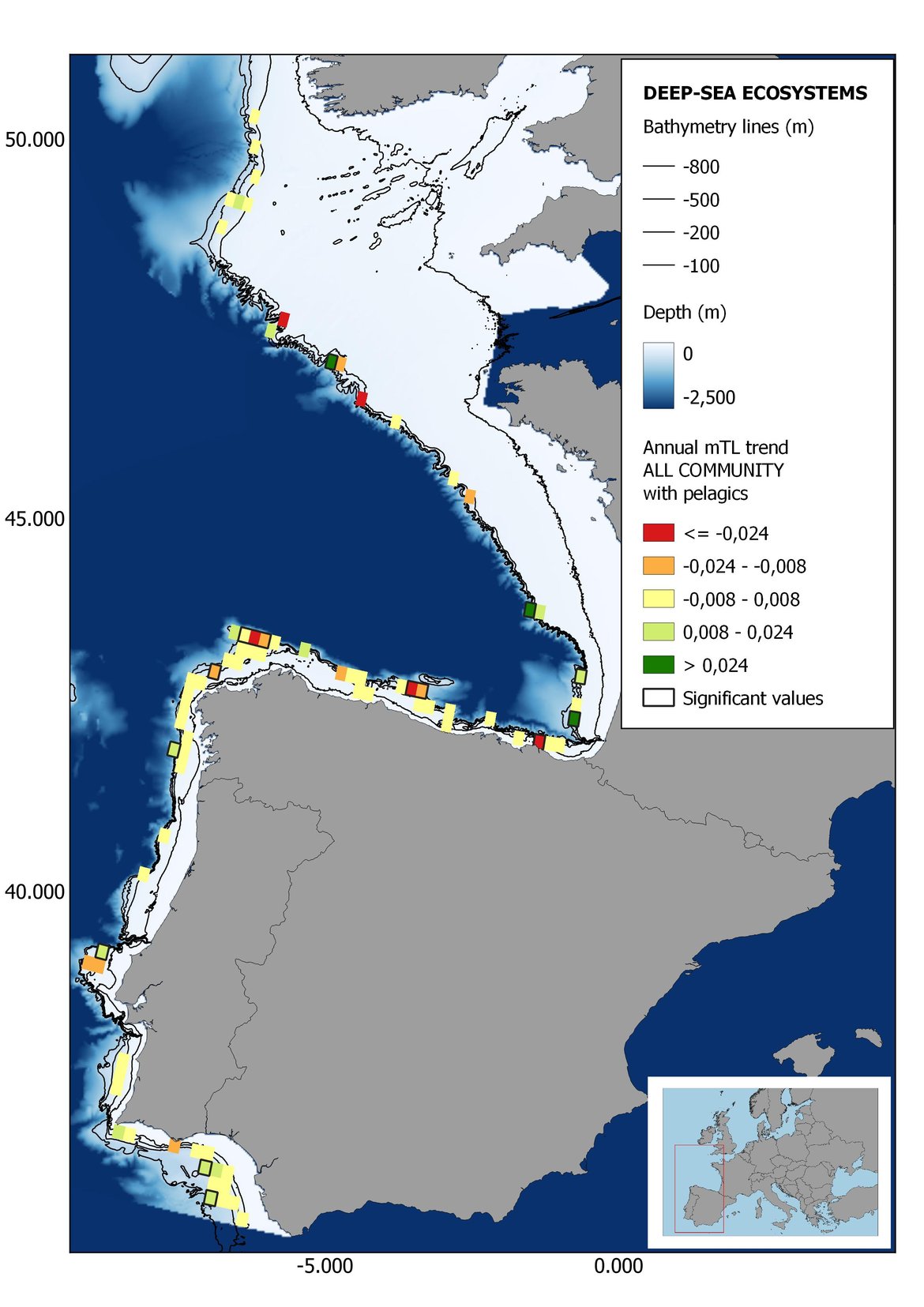
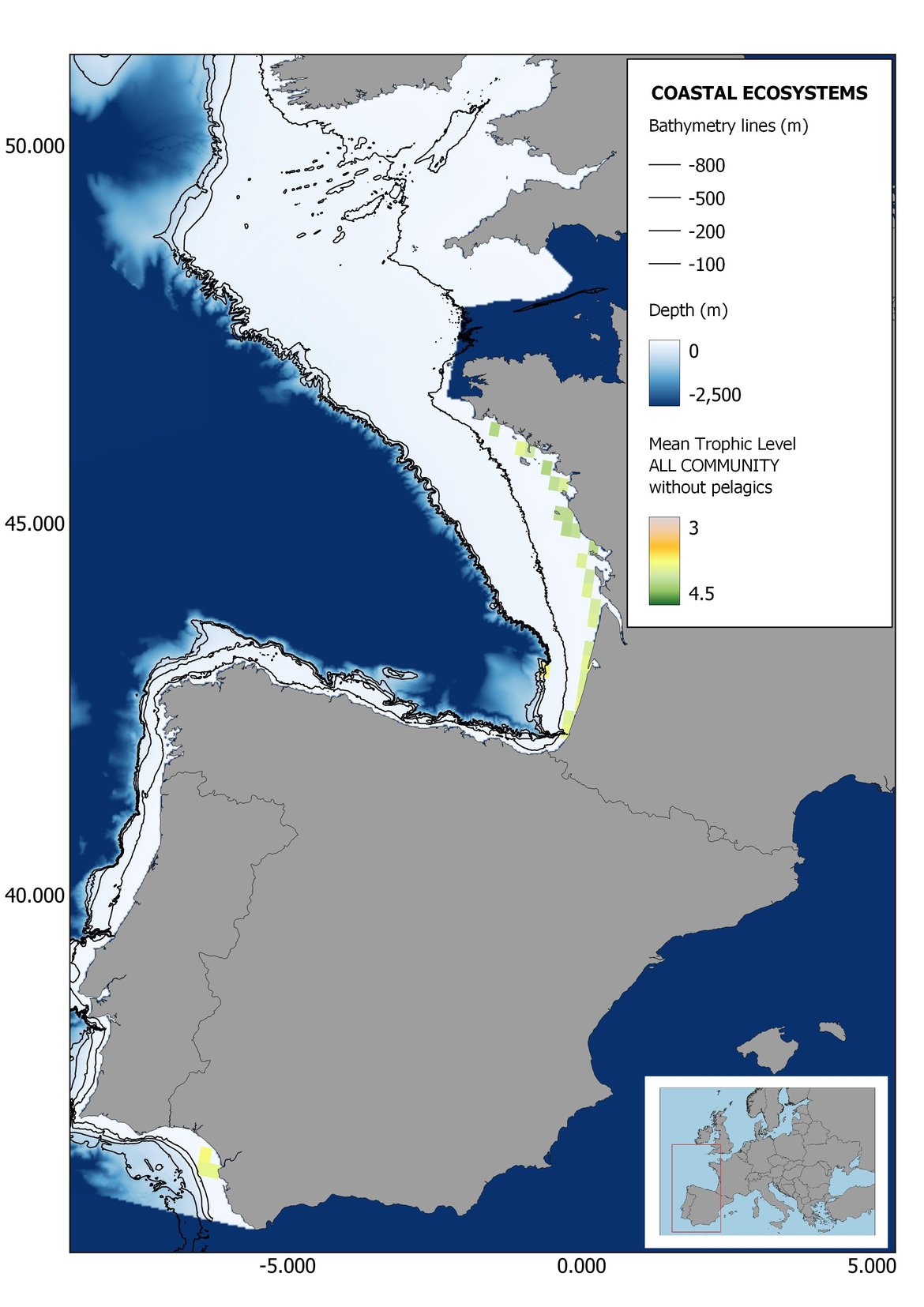
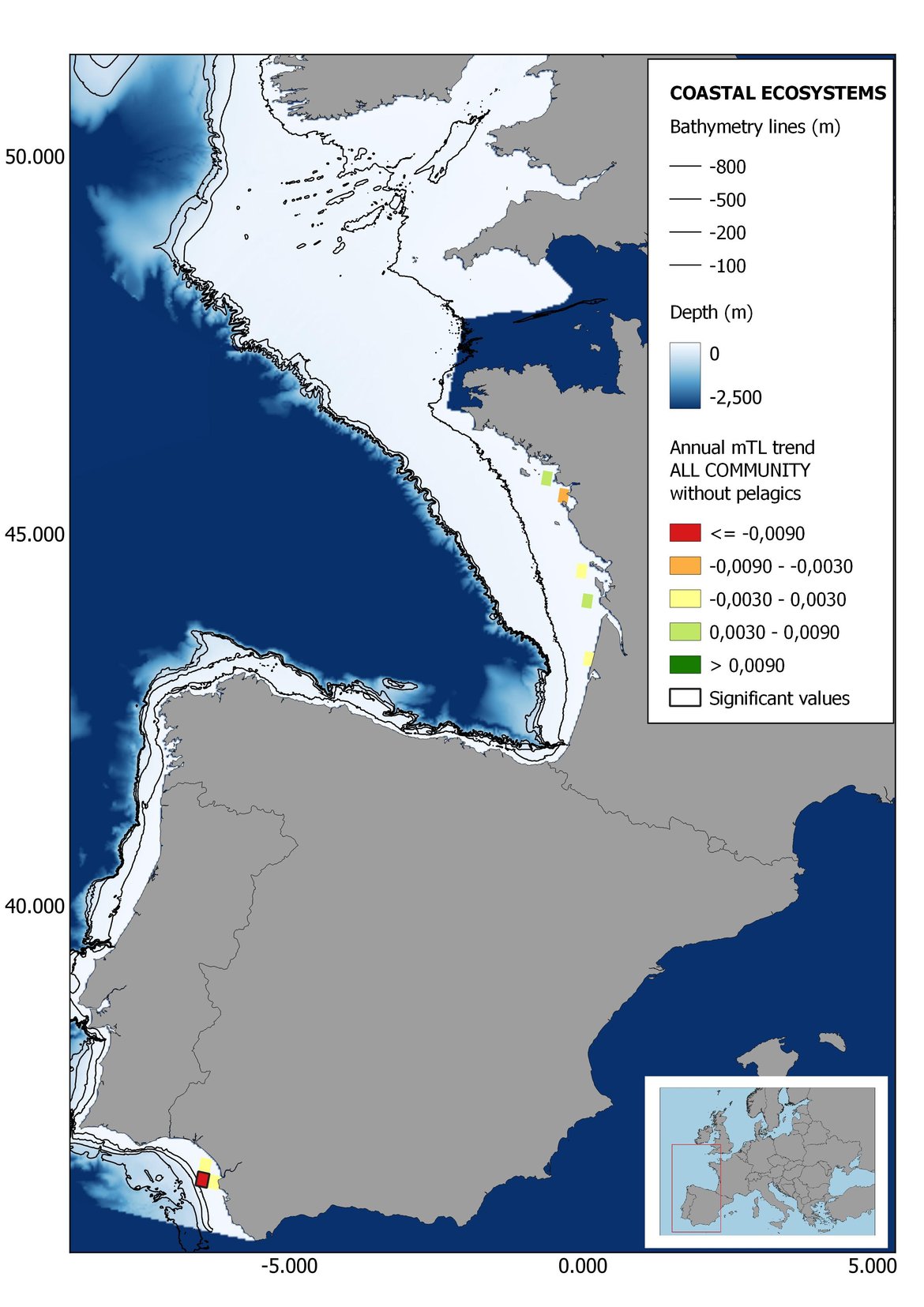
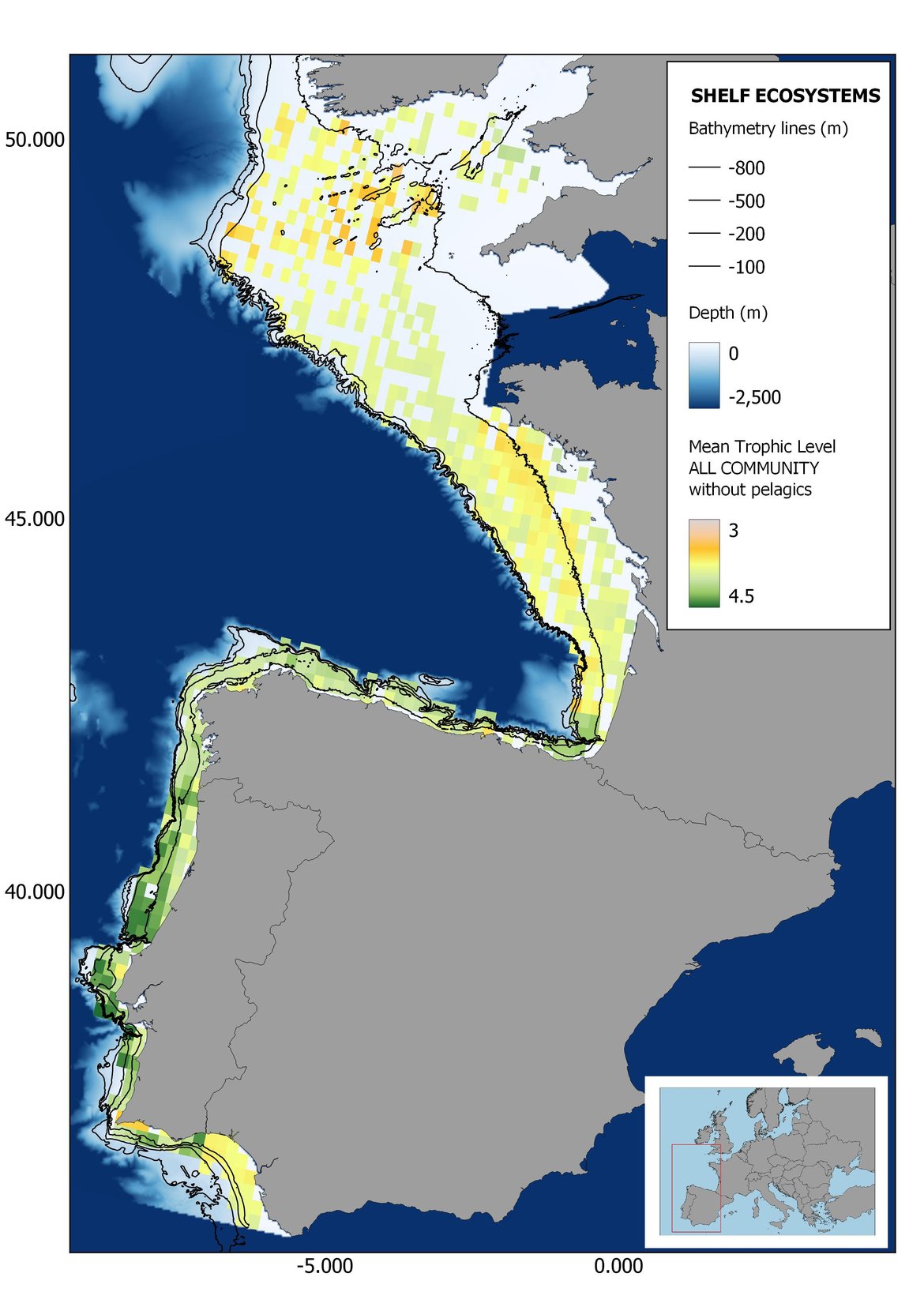
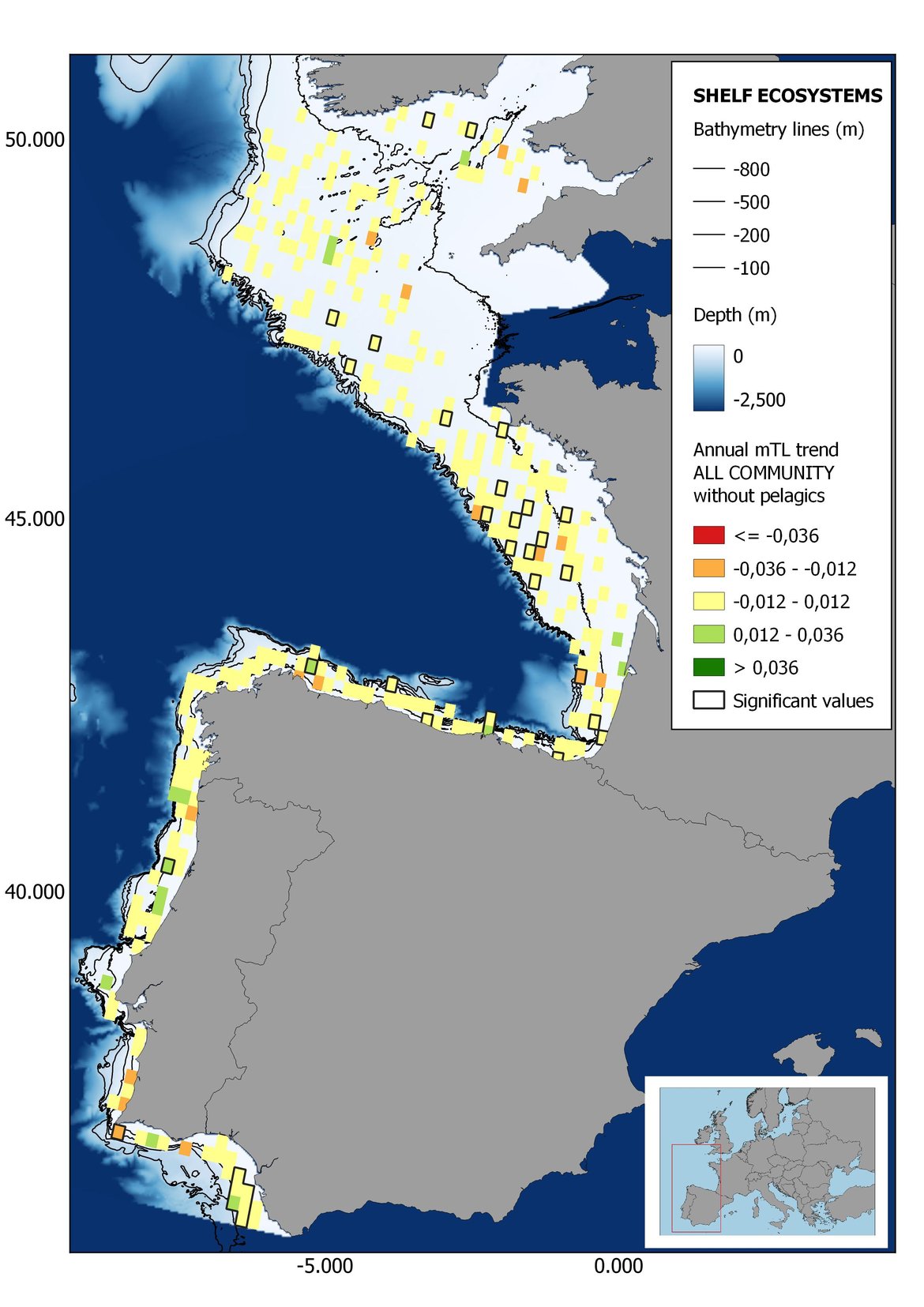
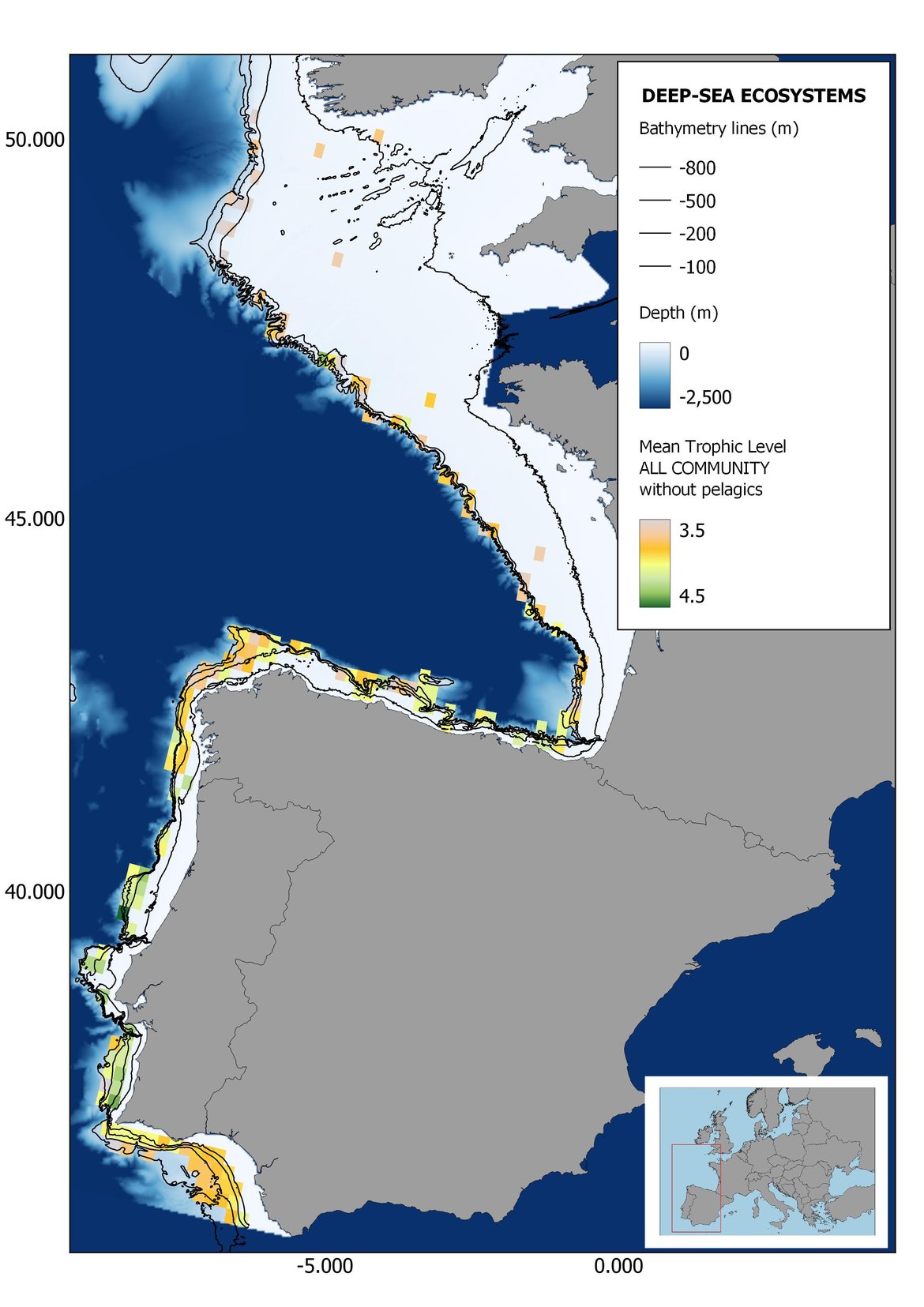
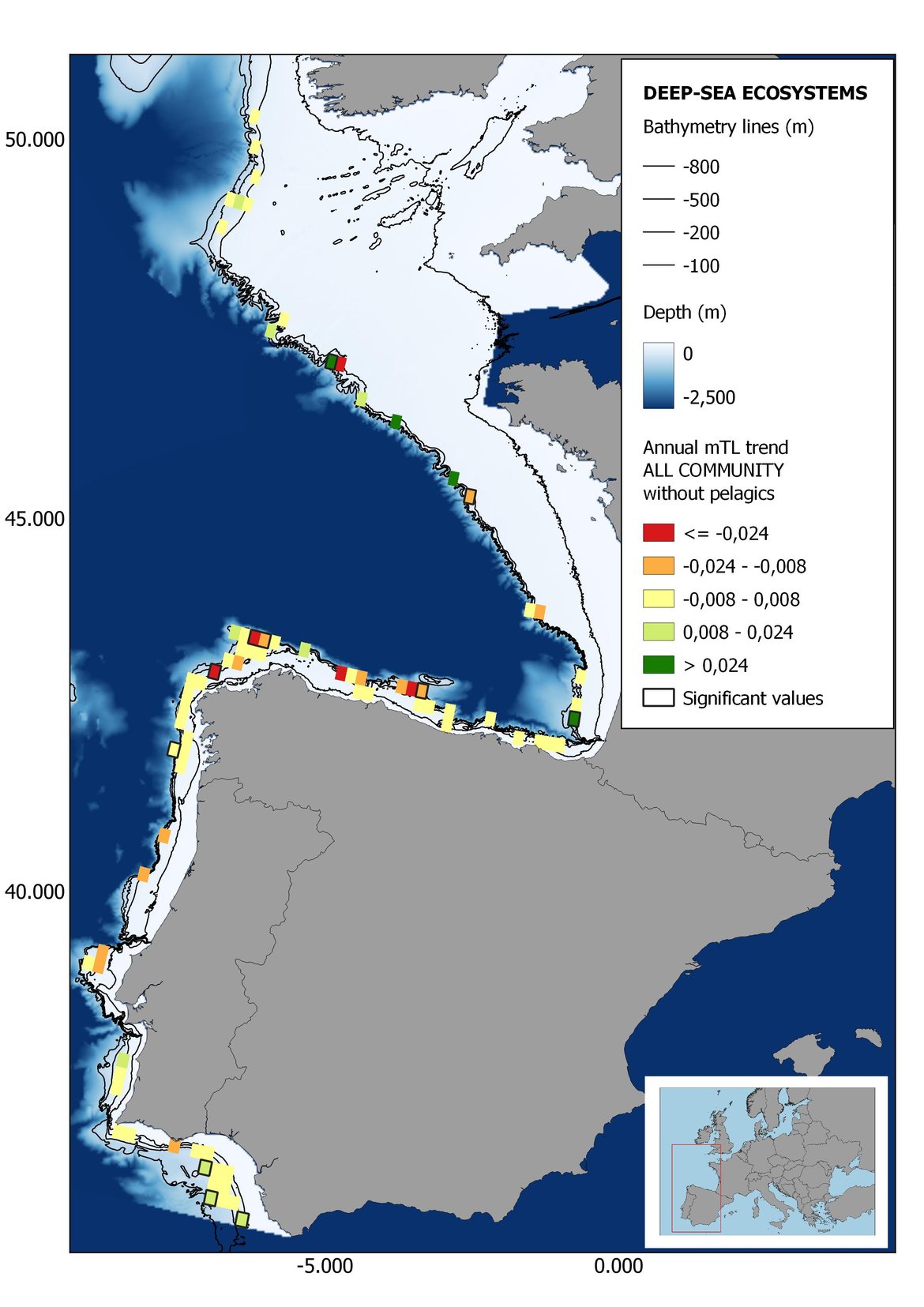
Figure 8: Maps showing the results of the spatio-temporal approach of all community: top 6 maps show mean Trophic Level by square, bottom 6 maps show trend of the MTL by square. Green = increasing trend, Orange-yellow = no trend, Red = decreasing trend. Significant trends are framed in bold.
The results of mapping the indicator show lower values of MTL in northern areas contrasting with higher MTL in Portuguese shelf ecosystem. When plotting the trends by square, small-scale temporal changes that were masked when the indicator was calculated by sub-division appeared. Looking at the results of Top predators, the northern area of the French and Portuguese shelf showed a patchy distribution of positive trends, while a concentration of decreasing trends can be seen in the north-western Iberian Peninsula (shelf and deep ecosystems). The decreasing trend in meso top predators observed in the temporal approach in the French shelf appeared to be related to high concentration of negative trends in southernmost area. Temporal and positive changes in meso-+top predators in North Iberian shelf are observed only in a few, scattered areas. The indicator including all community showed a high concentration of decreasing values in the northernmost part of the French shelf and North Iberian slope, while showing overall increasing trends in the northern Portuguese shelf and slope.
Table 1: Summary of the assessment of the different scenarios in the four OSPAR sub- divisions taking into account TL thresholds, depth strata and including/excluding pelagic species. Positive values indicate increasing trend; negative values indicate decreasing trend. Colours and categories are explained in Table b. The number in each cell represent the rate of change of the MTL across the whole historical series, calculated multiplying the slope of the linear by the total number of years of the time series.
| Top predators | Mesopredators + Top predators | All community | |||||||
|---|---|---|---|---|---|---|---|---|---|
| coast | shelf | deep | coast | shelf | deep | coast | shelf | deep | |
| France | |||||||||
| with pel | 0,03* | 0,03*** | 0,3 | 0,03 | 0,05*** | 0,02*** | -0,04 | -0,06** | -0,01 |
| without pel | 0,03* | 0,03*** | 0,03 | -0,04 | -0,03*** | 0,01 | 0,07 | -0,01 | -0,01 |
| North Iberian | |||||||||
| with pel | -0,03* | -0,04** | 0,06*** | 0,03* | 0,01 | -0,05 | |||
| without pel | -0,10*** | -0,10*** | 0,04** | 0,07*** | -0,01 | -0,06 | |||
| Portugal | |||||||||
| with pel | -0,05** | -0,04 | 0,06 | -0,01 | 0,02* | 0,07 | |||
| without pel | 0,05 | -0,04 | -0,06 | -0,03 | -0,09 | 0,02 | |||
| Gulf of Cadiz | |||||||||
| with pel | 0,02 | 0,03 | 0,15*** | -0,13* | 0,01 | 0,08* | -0,12* | -0,03 | 0,13*** |
| without pel | 0,07 | 0,03* | 0,14* | -0,10* | 0,15*** | 0,07* | -0,13** | 0,16*** | 0,16*** |
p-value for the statistical test of significance (linear model), *: p<0,05, **: p<0,01 ***: p<0,001.
In the present assessment the indicator MTL has been tested in the four sub-divisions within OSPAR Region IV: French, North Iberian, Portuguese and Gulf of Cadiz shelves. The compilation of TL values by sub-division for calculating MTL was possible for the main representative species in three out of four sub-divisions: French, North Iberian and Gulf of Cádiz continental shelves. The use of regional TLs is expected to improve the robustness of the indicator and reduce the uncertainty around the calculations.
French sub-division
Considering top predators alone (MTL4), the MTL indicator shows significant linear increasing trends in coastal and shelf areas, but no change in deeper waters. Similar responses are observed with and without pelagic species (Figure 2, Table 1). Considering meso + top predators (MTL3.25), the MTL indicator displayed contrasting responses: no significant trend was identified in coastal areas, but an increasing trend was displayed in continental shelf and slope which disappeared when the pelagic species were excluded from the community (Figure 2). No changes are observed when considering the whole community (MTL2) except for shelf ecosystems with pelagic, where a decreasing trend was detected.
Attending to the results of the spatio-temporal approach, the MTL of top predators (MTL4) showed no spatial differences with and without pelagic species (Figure 6). However, patchily distributed increasing trends were observed in the northernmost areas of the shelf ecosystem, around an area of lower mean MTL4 values in the Celtic Sea. The spatio-temporal analyses of meso + top predators MTL (MTL3.25), showed generally higher values on the shelf than on the slope (Figure 7). It’s noteworthy the decrease in MTL3.25 in these bathymetric ranges when excluding the pelagic species from the analyses, which is also associated to a change from mostly increasing trends to mostly decreasing trends in the annual values. (Figure 7). Regardless of the pelagic species, negative trends prevailed in the Southern Irish Sea (Figure 7). Attending to the whole community (MTL2), this change from mostly positive trends to mostly negative trends when excluding the pelagic species was also apparent (Figure 8) suggesting that the pelagic and demersal domains of the community are experiencing contrasting evolutions. When looking at the trends, no significant changes were observed in coastal areas at any of the MTL cut-offs (Figure 6, Figure 7 and Figure 8), but there was a clearly higher mean values in the northern area (Brittany and Pays de la Loire) than in the southern one (Nouvelle-Aquitaine) (Figure 6, Figure 7 and Figure 8). Overall, trends appeared heterogeneously and patchily distributed in all scenarios.
North Iberian sub-division
The indicator MTL4 (including top predators) showed moderate to strong decreasing trends in all scenarios: both depth strata (shelf and deep ecosystems), including and excluding pelagic species (Figure 3, Table 1). However, when including mesopredators, analysis revealed a significant positive trend in all scenarios. These contrasting trends could be detecting a signal of decline in biomass of top predators (some important commercial species) while an apparent improvement of mid trophic level consumers (mainly fish, elasmobranchs and cephalopods) in the last two decades. No changes have been observed when considering all community.
In the north Iberian sub-division, the MTL4 indicator (only top predators) showed higher values in the western part of the area in comparison to the inner Bay of Biscay (Figure 6), with almost no apparent differences including and excluding pelagic species. When looking at the trends of top predators, significant decreasing trends were observed in the westernmost area, more pronounced in shelf than in deep ecosystems (Figure 6), and also more pronounced when pelagic species were excluded from the analysis (Figure 6). Spatial differences in mean values and temporal changes were less evident when including mesopredators, although these contrasting trends (mostly negative at MTL4 and mostly positive at MTL3.25) were also observed at small scale. (Figure 6 and Figure 7). Taking into account all species in the community (MTL2) slope ecosystems displayed mostly decreasing trends in the indicator both including and excluding pelagic species, while shelf ecosystems showed mostly increasing trends in the indicator, regardless of the pelagic species (Figure 8).
Portuguese sub-division
In Portuguese waters, a moderate decline in MTL4 (top predators including pelagics) was observed in shelf ecosystems (Figure 4, Table 1). However, no changes are observed in the rest of scenarios, with the exception of a slight increase in the indicator including all community (MTL2 with pelagic) in shelf ecosystems (Table 1).
Higher values of MTL in Portuguese waters in comparison with other sub-divisions were evident when looking at the maps (Figure 6, Figure 7 and Figure 8). These results are consistently pronounced in almost all scenarios, and particularly evident in the central part of the Portuguese shelf and deep ecosystems. This sub-division showed little evidence of MTL changes at the local scale in any of the scenarios, with very few cells showing significant trends being scattered across the area (Figure 6, Figure 7 and Figure 8). These results at the local scale mostly agree with the temporal trends obtained when analysing the sub-division as a whole (Figure 4, Table 1), highlighting the greater stability/resilience of the Portuguese sub-division, in relation to the adjacent areas.
Gulf of Cadiz sub-division
Increasing trends in all scenarios excluding pelagic species were observed except for coastal areas (Figure 5 and Table 1) where a decline was evident when considering all community (MTL2) and meso + top predators (MTL3.25), but not for top predators only (MTL4). However, the results from coastal areas should be taken with caution due to the low number of hauls in comparison with shelf and deep ecosystems. It should be noted that the rate of change in MTL across the time series in many of the scenarios was one order of magnitude higher compared to other sub-divisions (Table 1).
Despite the low number of hauls analysed in coastal areas, the decreasing trends detected in the temporal approach can also be observed when mapping the indicator at small scale (Figure 6, Figure 7 and Figure 8). For top predators (MTL4) significant positive trends were detected in the southernmost part of the sub-division in shelf ecosystems, and generally over the continental slope (Figure 6). When mesopredators are included (MTL3.25), positive trends emerge in the southern part of the sub-division persist on the shelf, while areas with positive values on the slope have a scattered distribution (Figure 7). A few cells showed increasing trends when including all community (MTL2) in the scenarios where pelagic species were excluded (Figure 8).
In summary, analyses of groundfish data show an apparent improvement in top predators in the French continental shelf, in contrast to the decrease generally observed in North Iberian and Portuguese waters. In the Gulf of Cadiz however, there were signs of recovery in shelf and deep-sea ecosystems. When including mesopredators in the calculations, increasing trends were observed in most of the sub-divisions. The state of the marine food web displayed no apparent changes when analysing the entire community, with the exception of the Gulf of Cadiz, where contrasting trends were observed in the MTL depending on the depth strata considered (decrease versus increase in coastal and slope areas, respectively). These contrasting trends highlight the importance of focusing the assessment by sub-division, and also, the need to consider different bathymetric and ecological scenarios. Nevertheless, these findings need to be taken with caution as they are highly dependent on the length and source of the time series considered.
Conclusion
The Mean Trophic Level (MTL) in OSPAR Region IV showed contrasting trends in the overall food web structure, depending on the sub-division and the part of the ecosystem studied. In northern (French shelf) and southern areas (Gulf of Cadiz shelf) there were signs of improvements in the upper trophic levels, in contrast to the decline observed in top predators in the North Iberian and Portuguese shelf. However, when focusing on meso – and top predators the recovery is a common feature in most of the scenarios in the four sub-divisions. When taking into account the whole community, no changes were observed in most scenarios except for Gulf of Cadiz. The absence of variations in this scenario could be due to the masking effect of including many species with different feeding behaviours (fish, cephalopods and benthic invertebrates). An overall resilient food web due to persistent pressure over time could also explain the stable trends observed in the last two decades.
The use of trophic level values by sub-division provides a better view of community structure within the study region and strengthens the accuracy and robustness of this assessment. In addition, analysing the indicator with a spatio-temporal approach, the assessment provides complementary information of the evolution of the indicator at different scales, reinforcing the local effects on food web structure. Overall, examining the community as a whole (with and without pelagic species) as well as focusing on different trophic levels (top predators and mesopredators), allows revealing changes in food web structure that would remain hidden otherwise.
The Mean Trophic Level (MTL) in OSPAR Region IV showed an apparent sign of recovery when focusing on top predators (MTL4) in the northern and southernmost areas of the region (French continental and Gulf of Cadiz shelves). Nevertheless, when including mesopredators in the indicator (MTL3.25) an overall improvement was observed across sub-divisions. An increasing trend in the MTL indicator is considered to reflect decreasing impacts of fisheries on food webs, while a stable or declining trend needs to be further investigated in order to determine whether it can be attributed to resilience of the system or other possible causes. However, the contrasting trends observed in the different scenarios and sub-divisions support the idea that MTL should be assessed at sub-division level. Knowledge of the fisheries history in the region also helps to interpret these trends: the implementation of fisheries regulations may have decreased fishing pressure on top predators. Implementing fishing pressure data on the models will greatly help to investigate the direct response of the indicator. Additional information about the communities inhabiting the area and the environmental variability may be necessary to confirm the findings of this assessment. Furthermore, linking the MTL indicator to other biodiversity and food webs indicators (e.g., Feeding guilds) would also allow a more detailed examination of the impact of fisheries on the ecosystem.
Further work should be undertaken to improve the assessment of the state of the food web. Particularly important is the analysis of the interannual variability of TLs of species, which has been overlooked in the present assessment due to lack of appropriate data. Predator-prey interactions change over time, and therefore including these variations in the TL estimates will greatly improve the robustness of the models. In addition, as most species increase their trophic level along their ontogeny, mostly due to their larger size gape and increased motility, it would be more appropriate to estimate MTLs in the community based on both species and size distributions within the species. However, information of changes in TL with size are still unavailable for most of the species at the moment. Besides, incorporating additional datasets of other biological compartments (e.g., benthos, mammals, or birds) and / or from other sources (e.g., pelagic survey data) into the MTL indicator will enable a more holistic assessment of the area. Data on invertebrates (including cephalopods), which represent an important component of the biomass and play a key role as prey items with a fundamental role in the food web, should also be included in all sub-divisions.
An important effort has been made in the present assessment by performing the spatio-temporal approach, applying the indicator to finer geographic scales, identifying particularly impacted areas within those under high fishing pressure. However, it would be beneficial to further develop this indicator if more variables are included in the models (e.g., fishing pressure and climate variables), in order to explore the influence of anthropogenic pressure on MTL indicator trends, and how they affect the overall assessment.
Another issue is the difficulty of establishing reference conditions and assessment values. Indeed, the scientific survey time series used here started after the expansion of the fishing industry in this region and a period of high fishing pressure. The criteria used in the present assessment follow McQuatters-Gollop (2022), applying a combination of linear models and expert knowledge.
Knowledge Gaps
Using the present methodological approach, it is currently possible to apply the MTL indicator in other regions, whenever robust data on biomass and TLs of species are available. Lack of biomass data on benthos and invertebrates is a limitation to calculate the indicator for the whole community. Furthermore, detailed information on feeding interactions is strongly needed. Repeated dietary studies and stable isotope analyses implemented at (sub)regional scale and the computation of TL values would help fill in this gap in knowledge.
To improve the assessment of the state of the food web, further work on the MTL indicator include: i) to incorporate the interannual variability of the trophic level of species, ii) to incorporate variations of the trophic level of species with size, iii) to investigate the influence of various anthropogenic pressures on the indicator, iv) to improve the assessment criteria by defining thresholds using pressure-state curves.
In order to establish trophic relationships more accurately so that TL values can be determined, data are required on species feeding habits and their variations with ontogeny. Large-scale groundfish surveys (such as the Data Collection Framework, DCF) and pelagic surveys should aim at systematically collecting: i) as many species as possible in the food web; and ii) data for dietary analyses using stomach content and / or stable isotope analyses for the different size classes of a given species (in order to account for the ontogeny effect on species TL values). Comprehensive datasets on the feeding ecology of many of the key species in marine food webs are currently insufficient for the purposes of conducting this type of assessment, and this is especially the case for species at lower trophic levels.
The first step in filling these gaps in knowledge of food web structure and connectivity is to extend data collection on feeding ecology by sub-division (Carafa et al., 2007; Moloney et al., 2010; Rossberg et al., 2011). Analyses of stomach contents can provide accurate information on the species consumed as well as serving as a sampler of the smaller invertebrates and other benthic components not retained by the sampling gear. Stable isotope analyses indicate trophic level based on a longer feeding signature, and so provide a broader view of feeding strategy of a given species. Both approaches provide complementary information about food web functioning. Repeat dietary analyses also provide information on spatial and temporal variability in the trophic relationships of the species present in a given ecosystem and thus, of their TL values. Such data would be useful not only for strengthening the MTL indicator, but also for all trophic-level and food-web based indicators as well as for models.
Arroyo, N.L., Safi, G., Vouriot, P., López-López L., Niquil, N., Le Loc’h, F., Hattab T., Preciado I. 2019. Towards coherent GES assessments at sub-regional level: signs of fisheries expansion processes in the Bay of Biscay using an OSPAR food web indicator, the mean trophic level. ICES Journal of Marine Science. Available at: https://doi.org/10.1093/icesjms/fsz023.
Branch, T., Watson, R., Fulton, E., Jennings, S., McGillard, C., Pablico, G., Ricard, D., and Tracey, S. 2010. Trophic fingerprint of marine fisheries. Nature 468: 431-435.
Bundy, A., Shannon, L.J., Rochet, M.-J., Neira, S., Shin, Y.-J., Hill, L., Aydin, K., 2010. The good(ish), the bad and the ugly: a tripartite classification of ecosystem trends. ICES Journal of Marine Science 67, 745–768.
Carafa, R., Dueri, S., Zaldívar, and J.M. 2007. Linking terrestrial and aquatic ecosystems: Complexity, persistence and biodiversity in European food webs. In: EUR 22914 EN, Joint Research Centre.
Chassot, E., Rouyer, T., Trenkel, V.M., and Gascuel, D. 2008. Investigating trophic-level variability in Celtic Sea fish predators. Journal of Fish Biology 73: 763–781.
Chouvelon, T., Spitz, J., Caurant, F., Mèndez-Fernandez, P., Chappuis, A., Laugier, F., Le Goff, E., Bustamante, P., 2012. Revisiting the use of δ15N in meso-scale studies of marine food webs by considering spatio-temporal variations in stable isotopic signatures - The case of an open ecosystem: the Bay of Biscay (North-East Atlantic). Progress in Oceanography 101, 92–105. Available at: https://doi.org/10.1016/j.pocean.2012.01.004.
Coll, M., Shannon, L.J.; Kleisner, K.M.; Juan-Jordá, M.J.; Bundy, A.; Akoglu, A.G.; Banaru, D.; Boldt, J.L.; Borges, M.F.; Cook, A.; Diallo, I.; Fu, C.; Fox, C.; Gascuel, D.; Gurney, L.J.; Hattab, T.; Heymans, J.J.; Jouffre, D.; Knight, B.R.; Kucukavsar, S.; Large, S.I.; Lynam, C.; Machias, A.; Marshall, K.N.; Masski, H.; Ojaveer, H.; Piroddi, C.; Tam, J.; Thiao, D.; Thiaw, M.; Torres, M.A.; Travers-Trolet, M.; Tsagarakis, K.; Tuck, I.; van der Meeren, G.I.; Yemane, D.; Zador, S.G.; and Shin, Y.-J. 2016. Ecological indicators to capture the effects of fishing on biodiversity and conservation status of marine ecosystems. Ecological Indicators 60, 947–962.
Cury, P.M., Shannon, L.J., Roux, J.P., Daskalov, G.M., Jarre, A., Moloney, C.L., Pauly, D., 2005. Trophodynamic indicators for an ecosystem approach to fisheries. ICES Journal of Marine Science 62, 430–442
Essington, T.E., Beaudreau, A.H., and Wiedenmann, J. 2006. Fishing through marine food webs. Proceedings of the National Academy of Sciences USA 103: 3171−3175.
Heymans, J.J., Coll, M., Libralato, S., Morissette, L., Christensen, V., 2014. Global patterns in ecological indicators of marine food webs: a modeling approach. PLoS One 9 (4), e95845. Available at: https://doi.org/10.1371/journal.pone.0095845.
Lassalle, G., Lobry, J., Le Loc’h, F., Bustamante, P., Certain, G., Delmas, D., Dupuy, C., Hily, C., Labry, C., Le Pape, O., Marquis, E., Petitgas, P., Pusineri, C., Ridoux, V., Spitz, J., and Niquil, N. 2011. Lower trophic levels and detrital biomass control the Bay of Biscay continental shelf food web: implications for ecosystem management. Progress in Oceanography 91: 61–75
Lassalle, G., Chouvelon, T., Bustamante, P., and Niquil, N. 2014. An assessment of the trophic structure of the Bay of Biscay continental shelf food web: Comparing estimates derived from an ecosystem model and isotopic data. Progress in Oceanography 120: 205-2015.
Le Loc’h, F., Hily, C., and Grall, J. 2008. Benthic community and food web structure on the continental shelf of the Bay of Biscay (North Eastern Atlantic) revealed by stable isotopes analysis. Journal of Marine Systems 72: 17–34.
Link, J.S., Yemane, D., Shannon, L.J., Coll, M., Shin, Y.-J., Hill, L., Borges, M.F., 2010. Relating marine ecosystem indicators to fishing and environmental drivers: an elucidation of contrasting responses. ICES Journal of Marine Science 67, 787–795.
McQuatters-Gollop, A., Guérin, L., Arroyo, N.L., Aubert, A., Artigas, L.F., Bedford, J., Corcoran, E., Dierschke, V., Elliot, S.A.M., Geelhoed, S.C.V., Gilles, A., González-Irusta, J.M., Haelters, J., Johansen, M., Le Loc’h, F., Lynam, C.P., Niquil, N., Meakins, B., Mitchell, I., Padegimas, B., Pesch, R., Preciado, I., Rombouts, I., Safi, G., Schmitt, P., Schückel, U., Serrano, A., Stebbing, P., De la Torriente, A., Vina-Herbon, C. 2022. Assessing the state of marine biodiversity in the Northeast Atlantic. Ecological indicators. Available at: https://doi.org/10.1016/j.ecolind.2022.109148
Moloney, C.L., St John, M.A., Denman, K.L., Karl, D.M., Koster, F.W., Sundby, S., and Wilson, R.P. 2010. Weaving marine food webs from end to end under global change. Journal Of Marine System 84: 106-116.
Morato, T., Watson, R., Pitcher, T.J., and Pauly, D. 2006. Fishing down the deep. Fish 7: 24−34.
Pauly, D., and Watson, R. 2005. Background and interpretation of the ‘Marine Trophic Index’ as a measure of biodiversity. Philosophical Transactions of the Royal Society: Biological Sciences 360: 415-423.
Pauly, D., Christensen, V., Dalsgaard, J., Froese, R., and Torres Jr., F. 1998. Fishing down marine food webs. Science 279: 860 – 863.
Pauly, D., Hilborn, R., and Branch, A. 2013. Does catch reflect abundance? Nature 494: 303-306.
Pinnegar, J.K., Jennings, S., Brien, C.M.O., and Polunin, N.V.C. 2002. Long-term changes in the trophic level of the Celtic Sea fish community and fish market price distribution. Journal of Applied Ecology 39: 377-390.
Preciado I. Arroyo N.L., González-Irusta J.M., López-López L., Punzón A., Muñoz I., Serrano A. 2019 Small-scale spatial variations of trawling impact on food web structure. Ecological indicators 98: 442-452
R Core Team (2022). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at: https://www.R-project.org/.
Rossberg, A. G., Farnsworth, K. D., Satoh, K., and Pinnegar, J. K. 2011. Universal power-law diet partitioning by marine fish and squid with surprising stability-diversity implications. Proceedings of the Royal Society B 278 (1712): 1617–1625.
Shannon, L., Coll, M., Bundy, A., Gascuel, D., Heymans, J. J., Kleisner, K., Lynam, C. P., Piroddi, C., Tam, J., Travers-Trolet, M., and Shin, Y. 2014. Trophic level-based indicators to track fishing impacts across marine ecosystems. Marine Ecology Progress Series 512: 115-140.
Stergiou, K.I., and Tsikliras, A.C. 2011. Fishing down, fishing through and fishing up: fundamental process versus technical details. Marine Ecology Progress Series 441: 295−301.
Swartz, W., Sala, E., Tracey, S., Watson, R., and Pauly, D. 2010. The spatial expansion and ecological footprint of fisheries (1950 to present). PLoS ONE 5: e15143.
Vinagre, C., Salgado, J., Mendonca, V., Cabral, H., and Costa, M. 2012. Isotopes reveal fluctuation in trophic levels of estuarine organisms, in space and time. Journal of Sea Research 72:49-54.
Contributors
Authors: Preciado, I., López-López, L., Rabanal, I., Ortiz, J.J., Torres, M. Á., Muñoz, I., Iglesias, D., García Rebollo, J. M., Mendes, H., Le Loc’h, F., Garrido, S., Metaireau, H. and Serre, S.
Citation
Preciado, I., López-López, L., Rabanal, I., Ortiz, J.J., Torres, M. Á., Muñoz, I., Iglesias, D., García Rebollo, J. M., Mendes, H., Le Loc’h, F., Garrido, S., Metaireau, H. and Serre, S., 2023. Changes in Average Trophic Level of Marine Consumers. In: OSPAR, 2023: The 2023 Quality Status Report for the North-East Atlantic. OSPAR Commission, London. Available at: https://oap.ospar.org/en/ospar-assessments/quality-status-reports/qsr-2023/indicator-assessments/changes-avg-trophic-level-consumers-fw4
| Assessment type | Indicator Assessment |
|---|---|
| Summary Results | https://odims.ospar.org/en/submissions/ospar_change_mtl_consumer_msfd_2022_06/ |
| SDG Indicator | 14.2 By 2020, sustainably manage and protect marine and coastal ecosystems to avoid significant adverse impacts, including by strengthening their resilience, and take action for their restoration in order to achieve healthy and productive oceans |
| Thematic Activity | Biological Diversity and Ecosystems |
| Relevant OSPAR Documentation | OSPAR Agreement 2018‐08 CEMP Guidelines: Common Indicator in Region IV: Change in average trophic level of marine predators (FW4) |
| Linkage | https://www.ospar.org/documents?v=39002 |
| Date of publication | 2023-06-30 |
| Conditions applying to access and use | https://oap.ospar.org/en/data-policy/ |
| Data Snapshot | https://odims.ospar.org/en/submissions/ospar_change_mtl_consumer_snapsh_2022_06/ |
| Data Results | https://odims.ospar.org/en/submissions/ospar_change_mtl_consumer_result_2022_06/ |