

Status Assessment 2022 - European eel
Assessment of status | Distribution | Population/abundance | Condition | Previous OSPAR status assessment | Status (overall assessment) | |
|---|---|---|---|---|---|---|
Region | I | ←→ 2,5 | ←→1,3,5 | ?5 | ● | Poor |
| II | ←→2,5 | ←→1,3,5 | ?5 | ● | Poor | |
| III | ←→2,5 | ←→1,3,5 | ?5 | ● | Poor | |
| IV | ←→1 | ←→1,3,5 | ?5 | ● | Poor | |
| V | N/A | |||||
Explanation to table:
Distribution, Population size, Condition
Trends in status (since the assessment in the background document)
↓ decreasing trend or deterioration of the criterion assessed
↑ increasing trend or improvement in the criterion assessed
←→ no change observed in the criterion assessed
? trend unknown in the criterion assessed
Previous status assessment: If in QSR 2010 then enter Regions where species occurs ( ○) and has been recognised by OSPAR to be threatened and/or declining (● ) based on Chapter 10 Table 10.1 and Table 10.2 . If a more recent status assessment is available, then enter ‘poor’/’good’
Status*(overall assessment)
red – poor
green – good
? – status unknown
NA - Not Applicable
*applied to assessments of status of the feature or of a criterion, as defined by the assessment values used in the QSR 2023 or by expert judgement.
Key Pressure
↓ key pressures and human activities reducing
↑ key pressures and human activities increasing
←→ no change in key pressures and human activities
? Change in pressure and human activities uncertain
Threats or impacts (overall assessment)
red – significant threat or impact;
green–no evidence of a significant threat or impact
Blue cells – insufficient information available
NA – not applicable
1 – direct data driven
2 – indirect data driven
3 – third party assessment, close-geographic match
4 – third party assessment, partial-geographic match
5 – expert judgement
Assessment of key pressures | Barrier to species movement (Obstacles blocking access to upstream and downstream migrations) | Extraction of, or mortality/injury to, wild species (Water intake from hydropower turbines and pumping stations) | Input of other substances (synthetic/non-synthetic) (Effect on contaminants on reproductive potential) | Removal of target species (Legal fishing) | Removal of target species (Illegal Unreported Fishing (poaching) and Illegal trade) | Removal of non-target species (By-catch mortality) | Freshwater Habitat loss | Input or spread of non-indigenous species (Parasitic infection by nematode Anguillicola crassus) | Input or spread of non-indigenous species (non-indigenous predators); Loss of / change to natural biological communities (predation by natural predators) | Climate change | Threat or impact (conclusion based on the above rows) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
Region | I | ←→5 | ←→5 | ?5 | ↓3 | N/A | ←→5 | ←→5 | ←→1,5 | ↑5 | ↑1,5 | |
| II | ↓1,5 | ↑5 | ?5 | ↓3 | ?5 | ?5 | ↓5 | ←→1,5 | ↑5 | ↑1,5 | ||
| III | ↓1,5 | ↑5 | ?5 | ↓3 | ?5 | ?5 | ↓5 | ←→1,5 | ↑5 | ↑1,5 | ||
| IV | ←→5 | ↑5 | ?5 | ↓3 | ?5 | ?5 | ←→5 | ←→1,5 | ↑5 | ↑1,5 | ||
| V | N/A | |||||||||||
Confidence
High confidence in the status assessment of Population/abundance because of the quantitative data used, medium for the distribution and condition status.
High confidence in the assessment of legal fishery mortality threat because of the quantitative data used. Low confidence for others threats as quantitative data are lacking and the understanding of how these threats, individually or synergistically, contribute to decline in abundance is still poor.
Background Information
Year added to OSPAR List: 2008 (OSPAR Agreement 2008-6). The original evaluation of European eel against the Texel-Faial criteria referred to global/regional importance, sensitivity and decline criteria, with information also provided on threats.
( https://www.ospar.org/site/assets/files/1888/european_eel.pdf )
Summary extracted from the Case report (OSPAR, 2010).
- Global importance: As a conservative estimate, at least 80% (possibly 100%) of the larvae of European eel pass through the OSPAR Maritime Area, and at least 50% of the adult eels live in continental waters flowing into the area. Therefore, OSPAR Maritime Area is of global importance for Anguilla anguilla (OSPAR 2008).
- Regional importance: The European eel (Anguilla anguilla) is widely distributed in marine, coastal, brackish and freshwater habitats of Europe and occurs from the Atlantic coast of north Africa, in all of Europe (including Baltic Sea) and in the Mediterranean waters of Europe and northern Africa. In addition, the European eel also occurs in the Canary Islands, Madeira and the Azores Islands, and in Iceland. In Iceland, the eel situation is unique because it also harbours American eels (Anguilla rostrata) and natural hybrids of the two eel species (Albert et al., 2006; Avise et al., 1990). Unfortunately, there are insufficient data available to assess eel status in this particular area.
- Sensitivity: The European eel Anguilla anguilla has an unusual life history, making its sensitivity difficult to assess. Eels are long-lived and spawn only once in their lifetime (OSPAR, 2008). An analysis of the stock dynamics under different management regimes indicates that the recovery time for eel could be expected within 80 years or reducing fisheries to zero, whereas under an ultimately sustainable fishing regime of just 10% of the current rate of fishing mortality, recovery may take more than 200 years (Astrom & Dekker, 2007). There are however other models (Knights and Bonhommeau (unpublished data and analyses), in ICES (2009a) predict that a recovery of recruitment patterns to “normal” levels is possible within ten years under favourable climatic conditions. Other (non-natural) causes of mortality (including e.g., turbines) also need to be taken into account.
- Decline: The decline in glass eel recruitment was first noticed in 1985 (EIFAC, 1985). The prolonged decline in landings was mentioned as early as 1975 (ICES, 1976) but has received much less attention than that of recruitment. After a considerable drop in recruitment since the 1980s, glass eel abundance is still at a low level (Bornarel et al., 2018).
- Anthropogenic pressures and biological factors: Due to its unusual and complex life history, the reasons for the decline of the European eel are not fully understood. However, there are indications of linkages between the declines of eels and human activities, especially fisheries, construction of dams, weirs or embankments in rivers, chemical pollution, entrainment in water abstractions and loss / degradation of eel habitats.
Last status assessment: 2008. OSPAR (2010) concluded that Anguilla anguilla stock is facing an unprecedented level of decline. Anthropogenic factors (e.g. exploitation, habitat loss, contamination and transfer of parasites and diseases) as well as natural processes (e.g. climate change, predation) have likely contributed to the decline. It is hoped that 2007 EC Eel Regulation (Anon, 2007) should help address the fisheries threat. However, restrictions on fisheries alone will be insufficient, and management measures aimed at other anthropogenic impacts on habitat quality, quantity and accessibility will also be required. This concern has been corroborated by the current assessment.
Geographical Range and Distribution
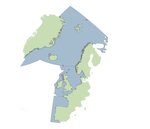
The European eel is a panmictic species, with a common spawning area in the Sargasso Sea. Eels undertake long migrations between the spawning areas and growth habitats. The latter extend to fresh, brackish and coastal waters of coastal countries in Europe, North Africa and Asian coasts of the Mediterranean and Black Sea (Adam, 1997; ICES, 2020b; Moriarty & Dekker, 1997). Oceanic migration routes are poorly understood. The northward branch of the Gulf Stream seems to be responsible for transporting most European eel larvae found north of about 40°N towards Europe (Miller et al., 2015). A recent tagging program demonstrated that silver eels in different locations in Europe were capable of long-distance migrations that would converge in the Azores region on a westerly route to the Sargasso Sea (Righton et al., 2016).
In continental waters, only physical obstacles seem to restrain the eel’s upstream distribution area. In the northern part (58-71°N) of its distribution range (Foldvik et al., 2019), as well as at the scales of France, Spain and Portugal bioregions analysed in the framework of the INTERREG SUDOANG ( https://sudoang.eu/fr/visuang/ ; Figure 2), there is no evidence that eel distribution has changed in recent years independently of potential local accessibility loss or gain.
Under conditions predicted for twenty-first century climate change, eel is expected to gain suitable basins mainly northward (around Baltic sea, Scandinavia and Arctic regions) (Lassalle & Rochard, 2009). On the opposite, a further increase in frequency and strength of droughts, induced by climate change and other anthropogenic pressures (reduction of flow regime, habitat loss) is likely to strongly impact the species distribution in the Iberian Peninsula (Costa et al., 2021).
Method of assessment: 2 and 5 for Artic waters, Greater North Sea and Celtic Sea; 1 for Bay of Biscay and Iberian coast (Region IV).
.")
Figure 1: Distribution area for A. anguilla, adapted from Adam (1997).
. Source: SUDOANG (https://aztidata.es/visuang/)")
Figure 2: Probability of presence of eel based on 21706 electrofishing data in SUDOE area (France, Spain and Portugal). Source: SUDOANG (https://aztidata.es/visuang/)
Population/Abundance
Indices of both glass and yellow eel recruitment declined strongly from 1980 to 2011. The latest glass eel recruitment in the “North Sea” index area, compared to that in 1960 to 1979, was 0,5% in 2020 (provisional) and 1,4% in 2019 (final). In the “Elsewhere Europe” index series it was 6.5% in 2020 (provisional) and 5,6% in 2019 (final), based on available data series. For the yellow eel data series, recruitment for 2019 was 17% (final) of the 1960 to 1979 level. Statistical analyses of the time series from 1980 to 2020 showed that glass eel recruitment remains at very low level (ICES, 2020a).
In a recent attempt, ICES WGEEL analysed short-term trends of silver eel abundances restricted to the period 2000 to 2019, using 17 time-series available in Europe (ICES, 2020b). While the majority of the time-series indicate a stable or declining trend of abundance, increases of abundance of silver eels have been observed in some river basins and no clear spatial pattern in the trends was observed (ICES, 2020b). This contrasted situation is likely to be related to different factors (environmental conditions, anthropogenic pressures, management practices) among river basins.
Method of assessment: 1, 3, 5 – Source: ICES (2020a; 2020b).
 glass eel recruitment for the continental “North Sea” and “Elsewhere Europe” series. The GLM was fitted to 52 time-series, comprising either pure glass eel or a mixture of glass + yellow eels (24 “North Sea” and 28 “Elsewhere Europe”). The predictions were scaled to the 1960 to 1979 average. Source: ICES (2020a).")
Figure 3: Indices, geometric mean of estimated (generalized linear model; GLM) glass eel recruitment for the continental “North Sea” and “Elsewhere Europe” series. The GLM was fitted to 52 time-series, comprising either pure glass eel or a mixture of glass + yellow eels (24 “North Sea” and 28 “Elsewhere Europe”). The predictions were scaled to the 1960 to 1979 average. Source: ICES (2020a).
 yellow eel recruitment trends for Europe. The GLM was fitted to 16 yellow eel time-series and scaled to the 1960-1979 average. Source: ICES (2020a).")
Figure 4: Estimated (GLM) yellow eel recruitment trends for Europe. The GLM was fitted to 16 yellow eel time-series and scaled to the 1960-1979 average. Source: ICES (2020a).
Condition
Since the mature oceanic key stage is not accessible, the ‘quality’ of silver eels leaving continental waters could be used as a proxy for how eels will perform in the future in terms of survival, migratory and reproductive success. Ecological and geographical clines, e.g. latitude and longitude, together with local environmental characteristics such as salinity, catchment size or food resources are known to influence silver eel traits. The generally recognized hypothesis is that there is (i) a positive and significant relation between age at silvering and latitude, and that (ii) size and thus fecundity increases with latitude (Vélez-Espino & Koops, 2010; Vollestad, 1992). Several laboratory experiments have shown that a fat content representing 12 to 13% of the body mass is required to fuel the trans-Atlantic migration, while a fat content estimated at 20% is thought to be necessary to ensure successful reproduction (Boetius & Boetius, 1980; Palstra & van den Thillart, 2010; Van Den Thillart et al., 2007). However, eels are particularly prone to bioaccumulation of (especially lipophilic) contaminants (Belpaire et al., 2016). Thus, levels of pollutants (PCBs) as well as the occurrence of infection by non-indigenous Anguillicola crassus parasite may have a significant effect upon migratory ability of silver eels.
Although variability in silver eel quality between rivers is observed (Bourillon et al., 2020), a reliable assessment of the stock quality is currently not possible, due to insufficient spatial and temporal data coverage (ICES, 2015).
Future prospect – Eel sex differentiation is highly influenced by the environment and particularly by density of congeners (Davey & Jellyman, 2005). Usually, males predominate in crowded areas such as downstream part of the watershed, while females prefer to inhabit the rivers upstream where densities are lowest (Geffroy & Bardonnet, 2015). In the context of eel decline, a lower density but higher proportion of larger females is expected (Davey & Jellyman, 2005; Poole et al., 2018).
Method of assessment: 5
Threats and Impacts
The eel is exposed to a multitude of pressures because of its diadromy and complex life history (Jacoby et al., 2015). For many of these pressures the impacts on the stock are difficult to assess and are largely unknown (ICES, 2020b). Many pressures often interact (e.g., blockage of upstream habitats might increase the density of the eel stock downstream, facilitating predation, fishing or disease transmission), which further complicates their assessment.
Obstacles blocking access to migration – Hundreds of years of modifications to water courses, e.g. dams, water abstraction structures or other barriers are major causes of reduced recruitment and spawner escapement. For instance, river fragmentation by dams is responsible for a 80% loss of historical range of the species in the Iberian Peninsula (Clavero & Hermoso, 2015). Much progress has been made in most Contracting Parties, as measures to improve connectivity are included in both EU Member States Eel Management Plans and river basins policies in the framework of EU Water Framework Directive (WFD). For instance, eel pass must be installed on all obstructions situated on designated ‘migratory fish’ rivers or on newly constructed barriers. With the increased ecological attention on re-establishing riverine connectivity, dam removal is also becoming an increasing river restoration measure in most Contracting Parties, particularly in the Greater North Sea (Region II) and Celtic Seas (Region III) (UK, Netherlands, Denmark, France, Sweden). However, river fragmentation is not easily reversed as more than 1,2 million instream barriers (with a mean density of 0,74 barriers per kilometer), 68% of which are structures less than two meters in height that are often overlooked, still exist in European rivers (Belletti et al., 2020).
Hydropower turbines and pumping stations – With their elongated morphology and poor burst swimming capabilities, eels are particularly vulnerable at screens and turbines (Bruijs & Durif, 2009; Righton et al., 2020). A wide range of technical and management solutions are available to enable mitigation of the impact of hydropower turbines and pumping stations: screening, deflection or diversion through passes, trap-and-transport, spillage of water when eels are migrating and the use of fish friendly designs of turbines and pumps (see ICES, 2019 for review). However, hydropower turbines and pumping stations are still a key sources of eel mortality (ICES, 2018) which is likely to have increased in recent years, stimulated by both recent technological developments and the need to produce renewable energy.
Effect of contaminants on reproductive potential – Since eels are benthic carnivores with a high fat content and long life span, they tend to accumulate higher amounts of persistent chemicals from water, food, and sediment than other species (Belpaire et al., 2016). There is evidence that some “historical” contaminants (PCB, DDT, pesticides, Lindane, etc.) have decreased in some place (Maes et al., 2008). However, the last Water Framework Directive evaluation pointed out that significant effort is required to meet chemical status across European surface waters as only 38% of surface waters are in good chemical status (European Commission, 2020). Moreover, emerging (e.g. toxic textile dyes or PFAS (per- and polyfluoralkyl substances); Belpaire et al. 2015; Teunen et al. 2021) and yet unknown hazardous substances may constitute additional threats for the species. Quantifying to what extent, and at what level, contaminants affect, singly or cumulatively, eel reproductive success is critical (Righton et al., 2020) but empirical data are still lacking.
Legal fishing – Glass eel landings show a sharp decline from 2 000 t in 1980 to around 59 t for the 2014 to 2018 period (ICES, 2020b). Commercial yellow and silver eel landings have declined from around 2 000 to 3 500 t in 2009 to around 2 691 t for the 2014 to 2018 period (ICES, 2020b). Threats from legal fishing appear to be decreasing due to a combination of local closures, reduction in European eel stocks and fishing effort. However, due to the critical status of the eel, the latest ICES Advice from 2021 proposes zero catch in all habitats for commercial and recreational fishing, including catches of glass eel for restocking and aquaculture (ICES, 2021).
Illegal Unreported Fishing and Illegal trade - There is an increasing amount of evidence that illegal, unreported and unregulated (IUU) fishing and illegal trade occur for glass and older eel stages (Crook, 2011; Richards et al., 2020; Shiraishi & Crook, 2015). Unfortunately, there are insufficient data available to quantify their effect on the total stock size or status at any level of certainty (ICES, 2020b) and to assess temporal trends at the scale of OSPAR Regions.
By-catch mortality – Most eels are caught in targeted fisheries in coastal waters, transitional (brackish) and freshwater. However, eels may be captured as by-catch in commercial and recreational fisheries (ICES, 2016). There is limited information on number of eels captured as by-catch, or on their survival when there are regulations requiring the release of eel captured in other fisheries (for instance by recreational angling).
Habitat loss – Aquatic habitat loss has been considerable throughout Europe during the last century, mainly because of wetland reclamation in coastal and estuarine environments but also due to floodplain drainage, dredging, etc. (Feunteun, 2002). In northern Europe where precipitation seems plentiful and river restoration programs are growing successfully (Peters et al., 2021), the threat from habitat loss appears to have decreased. Although now usually subject to more scrutiny from environmental regulators, there is still pressure in southwestern Europe (France, Spain and Portugal) to develop and alter floodplains and water courses for urban, industrial and agriculture development. Loss of habitat is still serious in this area where it is linked with drought and rapid economic development (Kettle et al., 2011). However, quantitative data are needed to fully understand the impact and trend of this pressure on the species (ICES, 2020b).
Anguillicola crassus – The nematode Anguillicola crassus has been shown to damage swim bladders and impair the swimming ability of infected eels (Palstra et al., 2007; Sjöberg et al., 2009). This parasite was introduced by human activities into European waters in the early 1980s and spread across the continent within 10 years (Kirk, 2003). Since then, A. crassus has become one on the most abundant parasites of the European eel (Gerard et al., 2013; Jakob et al., 2009) occurring in inland waters. Today, 60 to 100% of freshwater eels in eutrophic lakes are infected with A. crassus with a relatively stable trend of infection since 2010 (Acou, personal communication).
Natural and non-indigenous predators – In freshwater, several studies have indicated the significance of A. anguilla in the diets of various birds (Carpentier et al., 2009; e.g. sawbill ducks Mergus spp., grey heron Ardea cinerea, great cormorant Phalacrocorax carbo or osprey Pandion Haliaetus, Carss et al., 1999; Carss & Marquiss, 1997) or mammal predators such as by otters Lutra lutra (Carss et al., 1999). At sea, typical eel predators include marine mammals, sharks and large bony fishes (Battaglia et al., 2013; Beguer-Pon et al., 2012; Wahlberg et al., 2014). The recovery of, or increase in, predators such as the cormorants, seals or otters (Hansson et al., 2018) could increase the natural mortality rate in eel populations. Whilst a natural function of a healthy ecosystem, this may hinder the recovery of depleted species such as the European eel. In addition, the introduction and mismanagement of predators such as Wels catfish (Silurus glanis) for angling purpose could be an additional threat for the species in areas where this predator is established (principally OSPAR Region IV). However, the impact of this non-indigenous species on diadromous species including European eel is still unknown (Syväranta et al., 2009).
Climate change – Threats from climate change appear to impact all eel life-stages and all OSPAR Regions. Modifications of oceanic physical conditions appear to be impacting hatching, survival and drift of larvae towards European coasts (Drouineau et al., 2018). In river systems, changing patterns of precipitation and temperature are likely to make extreme events (floods, droughts) more common (Death et al., 2015), which could have serious effects on the quality of habitats and eel survival, especially in areas where other threats co-occur (e.g., drought and water abstraction). Regarding migration phenology, an earlier migration period of silver eel is expected in northern Europe due to increased river discharge in summer (Dankers & Feyen, 2008). Contrarily, drought periods in southern Europe are expected to extend into fall and delay migration (Arevalo et al., 2021).
Measures that address key pressures from human activities or conserve the species
The following list of measures was derived from the Contracting Parties implementation reporting, complemented by national expert additions.
- National Eel Management plans have been implemented by Belgium, Denmark, Finland, France, Germany, Ireland, Netherlands, Portugal, Sweden and UK.
- Legislation and measures (installation of fish passages, removals of dams and habitat restoration) have been introduced to restore habitat accessibility and extent of suitable habitats (in particular in estuaries, lowland rivers, floodplains and backwaters) in Belgium, Denmark, France, Ireland, Netherlands, Norway, Sweden and UK.
- Measures (deflection screens combined with bypass facility, fish-friendly technology, turbine closure during migration peaks, trap and transport) to reduce anthropogenic mortalities from hydropower turbines, pumping stations and other water intakes have been introduced in Belgium, Denmark, France, Ireland, Netherlands, Spain, Sweden and UK.
- Measures to further reduce emissions of hazardous substances are addressed by national legislation (PCB plan in France) and/or within the framework of EU Water Framework Directive in most Contracting Parties.
- Measures to increase the traceability of eels (especially for restocking of glass eels) traded from southern to northern Europe have been introduced.
- Measures to control and restrain illegal fisheries and illegal trade have been introduced by Belgium, Denmark, France, Netherlands, Portugal, Spain, Sweden. and the UK (it might be worth noting that as the UK is no longer a EU Member State, the Northern Ireland Protocol permits Northern Ireland to continue trading with the EU single market whilst trade between UK and the EU is not currently possible without a CITES permit). Fines are handed out and illegal catches, fishing equipment and used vessels can be confiscated. The European eel (Anguilla anguilla) has been listed in CITES Appendix II and in the EU implementation of CITES rules (Annex B to Council Regulation (EC) No 338/97; EU, 1996) since 13 March 2009. Since 2010, import and export of eel from the EU has been prohibited.
- Additional measures reported by experts: There are fishing regulations in Belgium, Denmark, France, Germany, Iceland, Portugal, Sweden and the UK.
Conclusion
Despite the measures taken by Contracting Parties to reduce the impact of fishing, hydropower, obstacles to migration and pollution, or to increase eel abundance through restocking, the status of European eel is still very poor in OSPAR Regions I to IV. Indeed, glass eel recruitment remains at a very low level. The threats include direct exploitation at different stages, blockages to migratory routes by dams and other structures, habitat degradation and loss, pollution, climate change, parasites and diseases.
There is insufficient spatial and temporal data coverage to quantitatively assess abundance of silver eels and their condition. Moreover, quantitative data are needed to fully understand the impacts of the numerous threats on the species. Therefore, it was not possible to assess trends and status within OSPAR Regions with a high level of confidence, except for fisheries exploitation. In line with ICES (2020b), it is of critical importance to better understand the impact of non-fisheries threats on the eel species. It is also important to improve knowledge on the relative importance of inland waters to coastal waters as habitats for eel on the recruitment of eel.
Knowledge gaps
- Concerns about the effects of eel stocking practices (e.g. spread of disease, illegal trade, stocking above dams and hydro barriers) and its effectiveness in contributing to increased silver eel production have been raised. There is evidence that translocated and stocked eel can contribute to yellow and silver eel production in recipient waters (Brämick et al., 2016; Dekker & Beaulaton, 2016; Matondo et al., 2021). However, there is also increasing evidence that some silver eels from the Baltic Sea display unexpected migration patterns (Tambets et al., 2021). Current knowledge about their orientation mechanisms implies that translocated eels will likely have difficulties in finding their spawning area (Durif et al., 2021). Whilst a local benefit may be apparent, an assessment of net benefit to the wider eel stock is unquantifiable and limited by the lack of knowledge on the spawning of any eel.
- Increased data collection and development of data collection infrastructure to better acknowledge the impacts of non-fisheries threats on eel status
- Studies to gain an overview of the prevalence, abundance and condition of the eels over its distribution area and to assess the effects of contaminants and disease factors on lipid metabolism, condition, migration capacities and reproduction;
- Further research on contribution of marine eels to spawning stocks
- Further research on oceanic migration routes,
- Further research on artificial breeding.
Method used
Main source of information:
- Assessment lead was coordinated by France, namely A. Acou (UMS OFB-CNRS-MNHN PatriNat / Management of Diadromous Fish in their Environment OFB-INRAE-Institut Agro-UPPA, France) and J. Raitif (UMS OFB-CNRS-MNHN PatriNat).
- Two online meetings (22 June 2021 and 20 September 2021) and emailing were needed with national experts in order complete data gathering, validate the methodology for assessment and review the final draft assessment.
- Experts involved in the assessment were:
Norway: O. Diserud, C. Durif
Sweden: W. Dekker
United Kingdom: T. Basic, C. Bean, A. Taylor, A. Walker
Belgium : C. Belpaire
France: L. Beaulaton, E. Feunteun, T. Trancart
Portugal : I. Domingos
Assessment is based upon:
a) Third party assessment (ICES WGEEL principally)
b) Literature studies
c) Direct or indirect data
d) Expert opinion of national experts
Adam, G. (1997). L’anguille européenne (Anguilla anguilla L. 1758): Dynamique de la sous-population du lac de Grand-Lieu en relation avec les facteurs environnementaux et anthropiques.
Albert, V., Jónsson, B., & Bernatchez, L. (2006). Natural hybrids in Atlantic eels (Anguilla anguilla, A. rostrata): Evidence for successful reproduction and fluctuating abundance in space and time. Molecular Ecology, 15(7), 1903–1916. https://doi.org/10.1111/j.1365-294X.2006.02917.x
Anon. (2007). Council Regulation (EC) No 1100/2007 of 18 September 2007 establishing measures for the recovery of the stock of European eel. Official Journal of the European Union, L248, 17–23.
Arevalo, E., Drouineau, H., Tétard, S., Durif, C. M. F., Diserud, O. H., Poole, W. R., & Maire, A. (2021). Joint temporal trends in river thermal and hydrological conditions can threaten the downstream migration of the critically endangered European eel. Scientific Reports, 11(1), 16927. https://doi.org/10.1038/s41598-021-96302-x
Astrom, M., & Dekker, W. (2007). When will the eel recover? A full life-cycle model. ICES Journal of Marine Science, 64, 1491–1498. https://doi.org/10.1093/icesjms/fsm122
Avise, J. C., Nelson, W. S., Arnold, J., Koehn, R. K., George, C., Williams, G. C., & Thorsteinsson, V. (1990). The evolutionary genetic status of Icelandic eels. Evolution, 44(5), 1254–1262. https://doi.org/10.2307/2409286
Battaglia, P., Andaloro, F., Consoli, P., Esposito, V., Malara, D., Musolino, S., Pedà, C., & Romeo, T. (2013). Feeding habits of the Atlantic bluefin tuna, Thunnus thynnus (L. 1758), in the central Mediterranean Sea (Strait of Messina). Helgoland Marine Research, 67(1), 97–107. https://doi.org/10.1007/s10152-012-0307-2
Beguer-Pon, M., Benchetrit, J., Castonguay, M., Aarestrup, K., Campana, S. E., Stokesbury, M. J. W., & Dodson, J. J. (2012). Shark Predation on Migrating Adult American Eels (Anguilla rostrata) in the Gulf of St. Lawrence. PLoS ONE, 7(10), e46830–e46830. https://doi.org/10.1371/journal.pone.0046830
Belletti, B., Garcia de Leaniz, C., Jones, J., Bizzi, S., Börger, L., Segura, G., Castelletti, A., van de Bund, W., Aarestrup, K., Barry, J., Belka, K., Berkhuysen, A., Birnie-Gauvin, K., Bussettini, M., Carolli, M., Consuegra, S., Dopico, E., Feierfeil, T., Fernández, S., … Zalewski, M. (2020). More than one million barriers fragment Europe’s rivers. Nature, 588(7838), 436–441. https://doi.org/10.1038/s41586-020-3005-2
Belpaire, C., Pujolar, J., Geeraerts, C., & Maes, G. (2016). Contaminants in Eels and their Role in the Collapse of the Eel Stocks. In Biology and Ecology of Anguillid Eels (pp. 225–250). https://doi.org/10.1201/b19925-13
Belpaire, C., Reyns, T., Geeraerts, C., & Van Loco, J. (2015). Toxic textile dyes accumulate in wild European eel Anguilla anguilla. Chemosphere, 138, 784–791. https://doi.org/10.1016/j.chemosphere.2015.08.007
Boetius, I., & Boetius, J. (1980). Experimental maturation of female silver eels, Anguilla anguilla. Estimates of fecundity and energy reserves for migration and spawning. Dana, 1, 1–28.
Bornarel, V., Lambert, P., Briand, C., Antunes, C., Belpaire, C., Ciccotti, E., Diaz, E., Diserud, O., Doherty, D., Domingos, I., Evans, D., de Graaf, M., O’Leary, C., Pedersen, M., Poole, R., Walker, A., Wickström, H., Beaulaton, L., & Drouineau, H. (2018). Modelling the recruitment of European eel (Anguilla anguilla) throughout its European range. ICES Journal of Marine Science, 75(2), 541–552. https://doi.org/10.1093/icesjms/fsx180
Bourillon, B., Acou, A., Trancart, T., Belpaire, C., Covaci, A., Bustamante, P., Faliex, E., Amilhat, E., Malarvannan, G., Virag, L., Aarestrup, K., Bervoets, L., Boisneau, C., Boulenger, C., Gargan, P., Becerra-Jurado, G., Lobón-Cerviá, J., Maes, G. E., Pedersen, M. I., … Feunteun, E. (2020). Assessment of the quality of European silver eels and tentative approach to trace the origin of contaminants – A European overview. Science of The Total Environment, 743, 140675. https://doi.org/10.1016/j.scitotenv.2020.140675
Brämick, U., Fladung, E., & Simon, J. (2016). Stocking is essential to meet the silver eel escapement target in a river system with currently low natural recruitment. ICES Journal of Marine Science, 73(1), 91–100. https://doi.org/10.1093/icesjms/fst034
Bruijs, M. C. M., & Durif, C. M. F. (2009). Silver eel migration and behaviour. In Spawning Migration of the European Eel: Reproduction index, a useful tool for conservation management (pp. 75–95). Springer. https://doi.org/10.1007/978-1-4020-9095-0_4
Carpentier, A., Marion, L., Paillisson, J. M., Acou, A., & Feunteun, E. (2009). Effects of commercial fishing and predation by cormorants on the Anguilla anguilla stock of a shallow eutrophic lake. Journal of Fish Biology, 74(9), 2132–2138. https://doi.org/10.1111/j.1095-8649.2009.02286.x
Carss, D. N., Elston, D. a, Nelson, K. C., & Kruuk, H. (1999). Spatial and temporal trends in unexploited yellow eel stocks in two shallow lakes and associated streams. Journal of Fish Biology, 55(3), 636–654. https://doi.org/10.1111/j.1095-8649.1999.tb00704.x
Carss, D. N., & Marquiss, M. (1997). The diet of cormorants Phalacrocorax carbo in scottish freshwaters in relation to feeding habitats and fisheries. Ekologia Polska, 45(1), 207–222.
Clavero, M., & Hermoso, V. (2015). Historical data to plan the recovery of the European eel. Journal of Applied Ecology, 52(4), 960–968. https://doi.org/10.1111/1365-2664.12446
Costa, M. J., Duarte, G., Segurado, P., & Branco, P. (2021). Major threats to European freshwater fish species. Science of The Total Environment, 797, 149105. https://doi.org/10.1016/j.scitotenv.2021.149105
Crook, V. (2011). Trade in European Eels: Recent developments under CITES and EU wildlife trade regulations. TRAFFIC Bulletin, 23(2), 71–74.
Dankers, R., & Feyen, L. (2008). Climate Change Impact on Flood Hazard in Europe: An Assessment Based on High-Resolution Climate Simulations. Journal of Geophysical Research, 113. https://doi.org/10.1029/2007JD009719
Davey, A. J. H., & Jellyman, D. J. (2005). Sex determination in freshwater eels and management options for manipulation of sex. Reviews in Fish Biology and Fisheries, 15, 37–52. https://doi.org/10.1007/s11160-005-7431-x
Death, R. G., Fuller, I. C., & Macklin, M. G. (2015). Resetting the river template: The potential for climate-related extreme floods to transform river geomorphology and ecology. Freshwater Biology, 60(12), 2477–2496. https://doi.org/10.1111/fwb.12639
Dekker, W., & Beaulaton, L. (2016). Faire mieux que la nature? The history of eel restocking in Europe. Environment and History, 22(2), 255–300. https://doi.org/10.3197/096734016X14574329314407
Durif, C. M. F., Stockhausen, H. H., Skiftesvik, A. B., Cresci, A., Nyqvist, D., & Browman, H. I. (2021). A unifying hypothesis for the spawning migrations of temperate anguillid eels. Fish and Fisheries, n/a(n/a). https://doi.org/10.1111/faf.12621
European Commission. (2020). Evaluation of Council Regulation (EC) No 1100/2007 of 18 September 2007 establishing measures for the recovery of the stock of European eel (Commission Staff Working Document SWD(2020) 35 final; p. 50 p. + annexes).
Feunteun, E. (2002). Management and restoration of European eel population (Anguilla anguilla): An impossible bargain. Ecological Engineering, 18(5), 575–591. https://doi.org/10.1016/S0925-8574(02)00021-6
Foldvik, A., Ulvan, E., Thorstad, E., & Hesthagen, T. (2019). Occurrence of the European eel in lakes in the northern part of its distribution area is limited to low-altitude coastal areas, likely due to topographical conditions. Boreal Environment Research, 24, 1–11.
Geffroy, B., & Bardonnet, A. (2015). Sex differentiation and sex determination in eels: Consequences for management. Fish and Fisheries, n/a-n/a. https://doi.org/10.1111/faf.12113
Gerard, C., Trancart, T., Amilhat, E., Faliex, E., Virag, L., Feunteun, E., & Acou, A. (2013). Influence of introduced vs. Native parasites on the body condition of migrant silver eels. Parasite, 20, 38. https://doi.org/10.1051/parasite/2013040
Hansson, S., Bergström, U., Bonsdorff, E., Härkönen, T., Jepsen, N., Kautsky, L., Lundström, K., Lunneryd, S.-G., Ovegård, M., Salmi, J., Sendek, D., & Vetemaa, M. (2018). Competition for the fish – fish extraction from the Baltic Sea by humans, aquatic mammals, and birds. ICES Journal of Marine Science, 75(3), 999–1008. https://doi.org/10.1093/icesjms/fsx207
ICES. (2015). Report of the Workshop of a Planning Group on the Monitoring of Eel Quality under the subject ‘Development of standardized and harmonized protocols for the estimation of eel quality’ (WKPGMEQ) (ICES CM 2014/SSGEF:14; p. 274 pp.).
ICES. (2016). Report of the Working Group on Eels (WGEEL), 15-22 September 2016, Cordoba, Spain (ICES CM 2016//ACOM:19; p. 107 pp.).
ICES. (2018). Report of the Joint EIFAAC/ICES/GFCM Working Group on Eels (WGEEL), 3-10 October 2017, Kavala, Greece. (ICES CM 2017/ACOM:15; p. 99 pp.).
ICES. (2019). Joint EIFAAC/ICES/GFCM Working Group on Eels (WGEEL) (ICES Scientific Reports 1:50; p. 177 pp.).
ICES. (2020a). European eel (Anguilla anguilla) throughout its natural range. In Report of the ICES Advisory Commitee, 2020, ICES Advice 2020, ele.2737.nea. https://doi.org/10.17895/ices.advice.5898
ICES. (2020b). Joint EIFAAC/ICES/GFCM Working Group on Eels (WGEEL). ICES Scientific Reports, 2(85), 223 pp. https://doi.org/10.17895/ICES.PUB.5982
ICES. (2021). European eel (Anguilla anguilla) throughout its natural range. In Report of the ICES Advisory Commitee, 2021. ICES Advice 2021, Ele.2737.Nea. https://doi.org/10.17895/ices.advice.7752
Jacoby, D. M. P., Casselman, J. M., Crook, V., DeLucia, M.-B., Ahn, H., Kaifu, K., Kurwie, T., Sasal, P., Silfvergrip, A. M. C., Smith, K. G., Uchida, K., Walker, A. M., & Gollock, M. J. (2015). Synergistic patterns of threat and the challenges facing global anguillid eel conservation. Global Ecology and Conservation, 4, 321–333. https://doi.org/10.1016/j.gecco.2015.07.009
Jakob, E., Hanel, R., Klimpel, S., & Zumholz, K. (2009). Salinity dependence of parasite infestation in the European eel Anguilla anguilla in northern Germany. ICES Journal of Marine Science, 66(2), 358–366. https://doi.org/10.1093/icesjms/fsn160
Kettle, A. J., Vollestad, L. A., & Wibig, J. (2011). Where once the eel and the elephant were together: Decline of the European eel because of changing hydrology in southwest Europe and northwest Africa? Fish and Fisheries, 12(4), 380–411. https://doi.org/10.1111/j.1467-2979.2010.00400.x
Kirk, R. S. (2003). The impact of Anguillicola crassus on European eels. Fisheries Management and Ecology, 10(6), 385–394. https://doi.org/10.1111/j.1365-2400.2003.00355.x
Lassalle, G., & Rochard, E. (2009). Impact of twenty-first century climate change on diadromous fish spread over Europe, North Africa and the Middle East. Global Change Biology, 15(5), 1072–1089. https://doi.org/10.1111/j.1365-2486.2008.01794.x
Maes, J., Belpaire, C., & Goemans, G. (2008). Spatial variations and temporal trends between 1994 and 2005 in polychlorinated biphenyls, organochlorine pesticides and heavy metals in European eel (Anguilla anguilla L.) in Flanders, Belgium. Environmental Pollution, 153(1), 223–237. https://doi.org/10.1016/j.envpol.2007.07.021
Matondo, B. N., Delrez, N., Bardonnet, A., Vanderplasschen, A., Joaquim-Justo, C., Rives, J., Benitez, J.-P., Dierckx, A., Séleck, E., Rollin, X., & Ovidio, M. (2021). A complete check-up of European eel after eight years of restocking in an upland river: Trends in growth, lipid content, sex ratio and health status. Science of The Total Environment, 151020. https://doi.org/10.1016/j.scitotenv.2021.151020
Miller, M. J., Bonhommeau, S., Munk, P., Castonguay, M., Hanel, R., & Mccleave, J. D. (2015). A century of research on the larval distributions of the Atlantic eels: A re-examination of the data. Biological Reviews, 90(4), 1035–1064. https://doi.org/10.1111/brv.12144
Moriarty, C., & Dekker, W. (1997). Management of the European eel. Fisheries Bulletin, 15, 110–110.
OSPAR. (2010). Background Document for European eel Anguilla anguilla (OSPAR Commision Report, p. 24 p. + annex).
Palstra, A. P., Heppener, D. F. M., van Ginneken, V. J. T., Székely, C., & van den Thillart, G. E. E. J. M. (2007). Swimming performance of silver eels is severely impaired by the swim-bladder parasite Anguillicola crassus. Journal of Experimental Marine Biology and Ecology, 352(1), 244–256. https://doi.org/10.1016/j.jembe.2007.08.003
Palstra, A. P., & van den Thillart, G. E. E. J. M. (2010). Swimming physiology of European silver eels (Anguilla anguilla L.): Energetic costs and effects on sexual maturation and reproduction. Fish Physiology and Biochemistry, 36(3), 297–322. https://doi.org/10.1007/s10695-010-9397-4
Peters, B., Buuren, M., Herik, K., Daalder, M., Tempels, B., Rijke, J., & Pedroli, B. (2021). The Smart Rivers approach: Spatial quality in flood protection and floodplain restoration projects based on river DNA. WIREs Water, 8(3). https://doi.org/10.1002/wat2.1511
Poole, W. R., Diserud, O. H., Thorstad, E. B., Durif, C. M., Dolan, C., Sandlund, O. T., Bergesen, K., Rogan, G., Kelly, S., & Vøllestad, L. A. (2018). Long-term variation in numbers and biomass of silver eels being produced in two European river systems. ICES Journal of Marine Science, 75, 1627–1637. https://doi.org/10.1093/icesjms/fsy053
Richards, J. L., Sheng, V., Yi, C. W., Ying, C. L., Ting, N. S., Sadovy, Y., & Baker, D. (2020). Prevalence of critically endangered European eel (Anguilla anguilla) in Hong Kong supermarkets. Science Advances, 6(10), eaay0317. https://doi.org/10.1126/sciadv.aay0317
Righton, D., Piper, A., Aarestrup, K., Amilhat, E., Belpaire, C., Casselman, J., Castonguay, M., Díaz, E., Dörner, H., Faliex, E., Feunteun, E., Fukuda, N., Hanel, R., Hanzen, C., Jellyman, D., Kaifu, K., McCarthy, K., Miller, M. J., Pratt, T., … Gollock, M. (2020). Important questions to progress science and sustainable management of anguillid eels. Fish and Fisheries, n/a(n/a). https://doi.org/10.1111/faf.12549
Righton, D., Westerberg, H., Feunteun, E., Økland, F., Gargan, P., Amilhat, E., Metcalfe, J., Lobon-Cervia, J., Sjöberg, N., Simon, J., Anthony, A., Marisa, V., Alan, W., Thomas, T., Uwe, B., & Kim, A. (2016). Empirical observations of the spawning migration of European eels: The long and dangerous road to the Sargasso Sea. Science Advances, 2(10), e1501694–e1501694. https://doi.org/10.1126/sciadv.1501694
Shiraishi, H., & Crook, V. (2015). An analysis of Anguilla production, trade and consumption in East Asia (p. 53).
Sjöberg, N. B., Petersson, E., Wickström, H., & Hansson, S. (2009). Effects of the swimbladder parasite Anguillicola crassus on the migration of European silver eels Anguilla anguilla in the Baltic Sea. Journal of Fish Biology, 74(9), 2158–2170. https://doi.org/10.1111/j.1095-8649.2009.02296.x
Syväranta, J., Cucherousset, J., Kopp, D., Crivelli, A., Céréghino, R., & Santoul, F. (2009). Dietary breadth and trophic position of introduced European catfish Silurus glanis in the River Tarn (Garonne River basin), Southwest France. Aquatic Biology, 8(2), 137–144. https://doi.org/10.3354/ab00220
Tambets, M., Kärgenberg, E., Järvalt, A., Økland, F., Kristensen, M. L., Koed, A., & Bernotas, P. (2021). Migrating silver eels return from the sea to the river of origin after a false start. Biology Letters, 17(9), 20210346. https://doi.org/10.1098/rsbl.2021.0346
Teunen, L., Bervoets, L., Belpaire, C., De Jonge, M., & Groffen, T. (2021). PFAS accumulation in indigenous and translocated aquatic organisms from Belgium, with translation to human and ecological health risk. Environmental Sciences Europe, 33(1), 39. https://doi.org/10.1186/s12302-021-00477-z
Van Den Thillart, G., Palstra, A., & Van Ginneken, V. (2007). Simulated migration of European silver eel; swim capacity and cost of transport. Journal of Marine Science and Technology, 15(SPEC. ISS.), 1–16.
Vélez-Espino, L. A., & Koops, M. A. (2010). A synthesis of the ecological processes influencing variation in life history and movement patterns of American eel: Towards a global assessment. Reviews in Fish Biology and Fisheries, 20(2), 1–24. https://doi.org/10.1007/s11160-009-9127-0
Vollestad, L. A. (1992). Geographic variation in age and length at metamorphosis of maturing European eel: Environmental effects and phenotypic plasticity. Journal of Animal Ecology, 61(1), 41–48.
Wahlberg, M., Westerberg, H., Aarestrup, K., Feunteun, E., Gargan, P., & Righton, D. (2014). Evidence of marine mammal predation of the European eel (Anguilla anguilla L.) on its marine migration. Deep Sea Research Part I: Oceanographic Research Papers, 86, 32–38. https://doi.org/10.1016/j.dsr.2014.01.003
Sheet reference:
BDC2022/European eel